

Out-of-register β-sheets suggest a pathway to toxic amyloid aggregates
- PMID: 23213214
- PMCID: PMC3529048
- DOI: 10.1073/pnas.1218792109
Out-of-register β-sheets suggest a pathway to toxic amyloid aggregates
Abstract
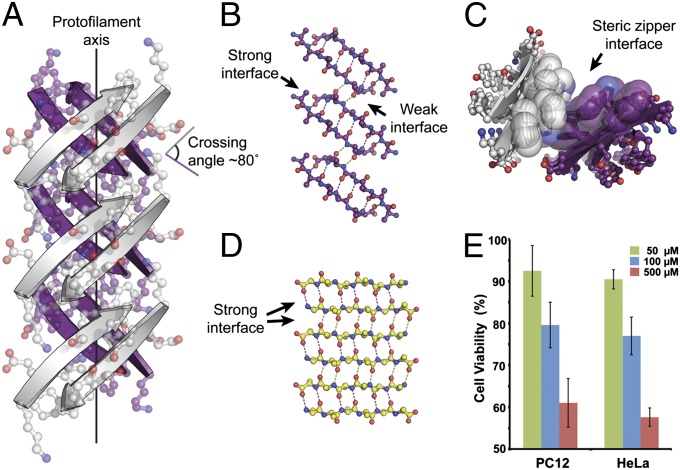
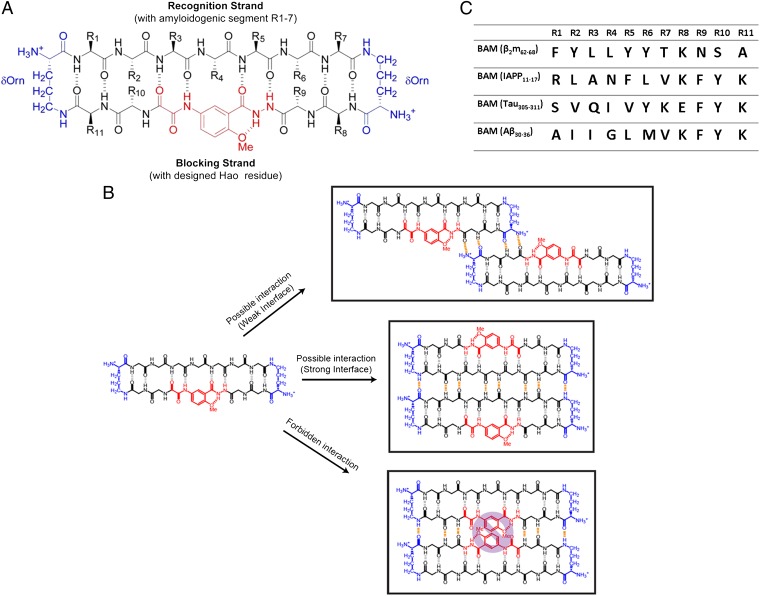
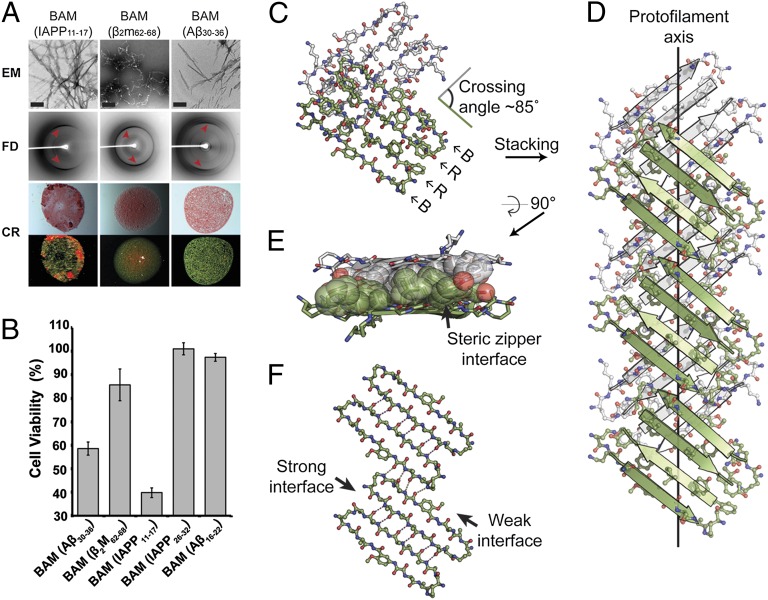
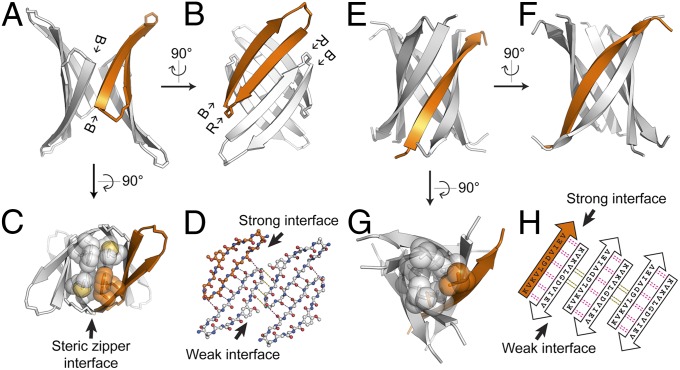
Although aberrant protein aggregation has been conclusively linked to dozens of devastating amyloid diseases, scientists remain puzzled about the molecular features that render amyloid fibrils or small oligomers toxic. Here, we report a previously unobserved type of amyloid fibril that tests as cytotoxic: one in which the strands of the contributing β-sheets are out of register. In all amyloid fibrils previously characterized at the molecular level, only in-register β-sheets have been observed, in which each strand makes its full complement of hydrogen bonds with the strands above and below it in the fibril. In out-of-register sheets, strands are sheared relative to one another, leaving dangling hydrogen bonds. Based on this finding, we designed out-of-register β-sheet amyloid mimics, which form both cylindrin-like oligomers and fibrils, and these mimics are cytotoxic. Structural and energetic considerations suggest that out-of-register fibrils can readily convert to toxic cylindrins. We propose that out-of-register β-sheets and their related cylindrins are part of a toxic amyloid pathway, which is distinct from the more energetically favored in-register amyloid pathway.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
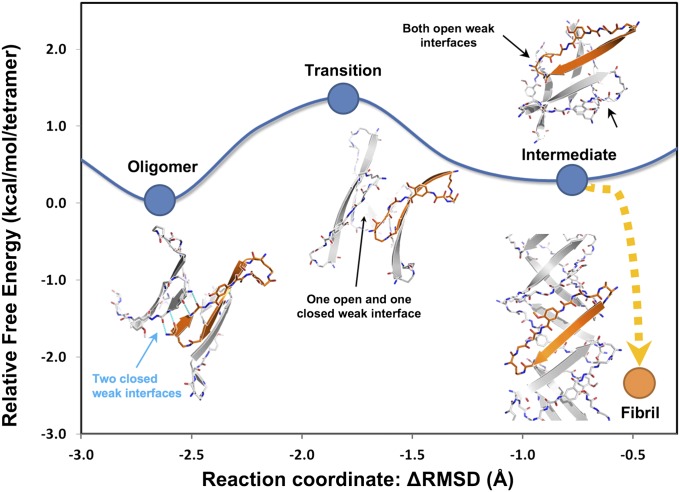
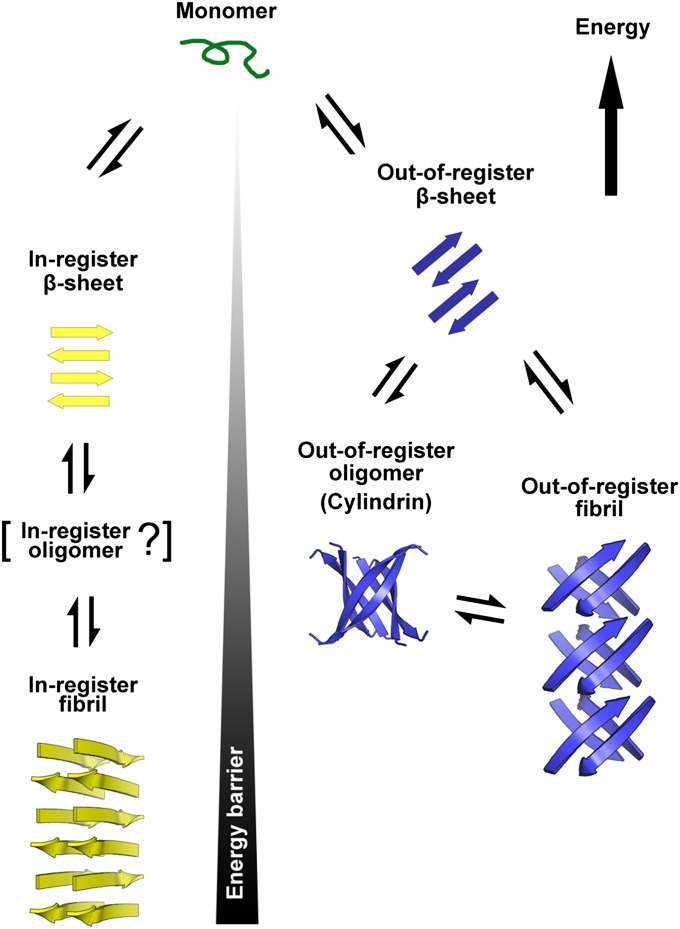
References
-
- Chiti F, Dobson CM. Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem. 2006;75:333–366. - PubMed
-
- Härd T, Lendel C. Inhibition of amyloid formation. J Mol Biol. 2012;421(4–5):441–465. - PubMed
-
- Eichner T, Radford SE. A diversity of assembly mechanisms of a generic amyloid fold. Mol Cell. 2011;43(1):8–18. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
