

Arabidopsis lateral organ boundaries negatively regulates brassinosteroid accumulation to limit growth in organ boundaries
- PMID: 23213252
- PMCID: PMC3529045
- DOI: 10.1073/pnas.1210789109
Arabidopsis lateral organ boundaries negatively regulates brassinosteroid accumulation to limit growth in organ boundaries
Abstract
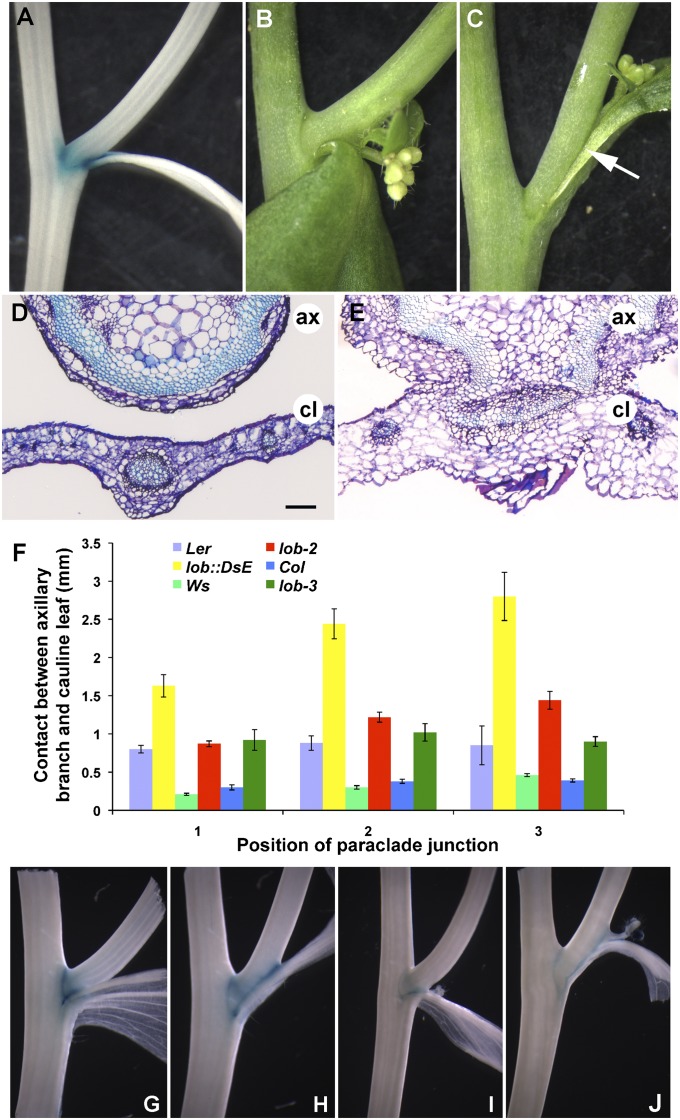
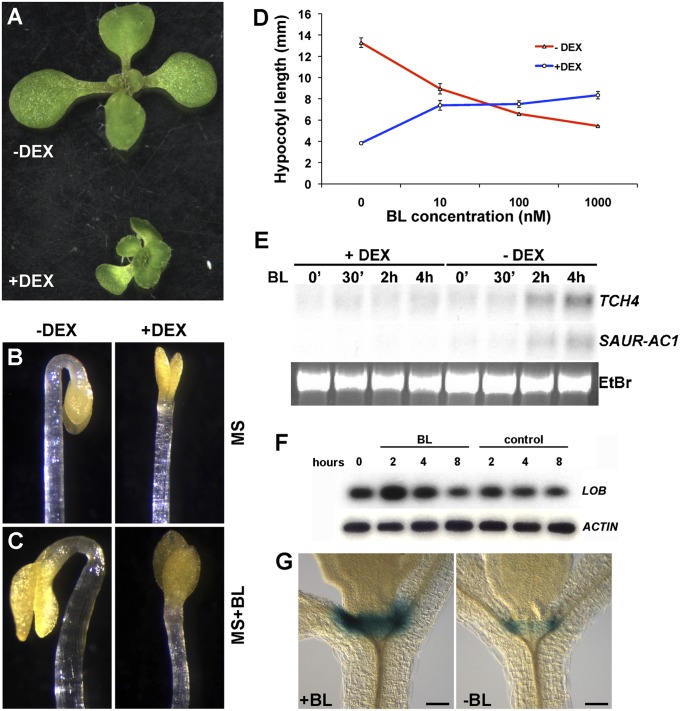
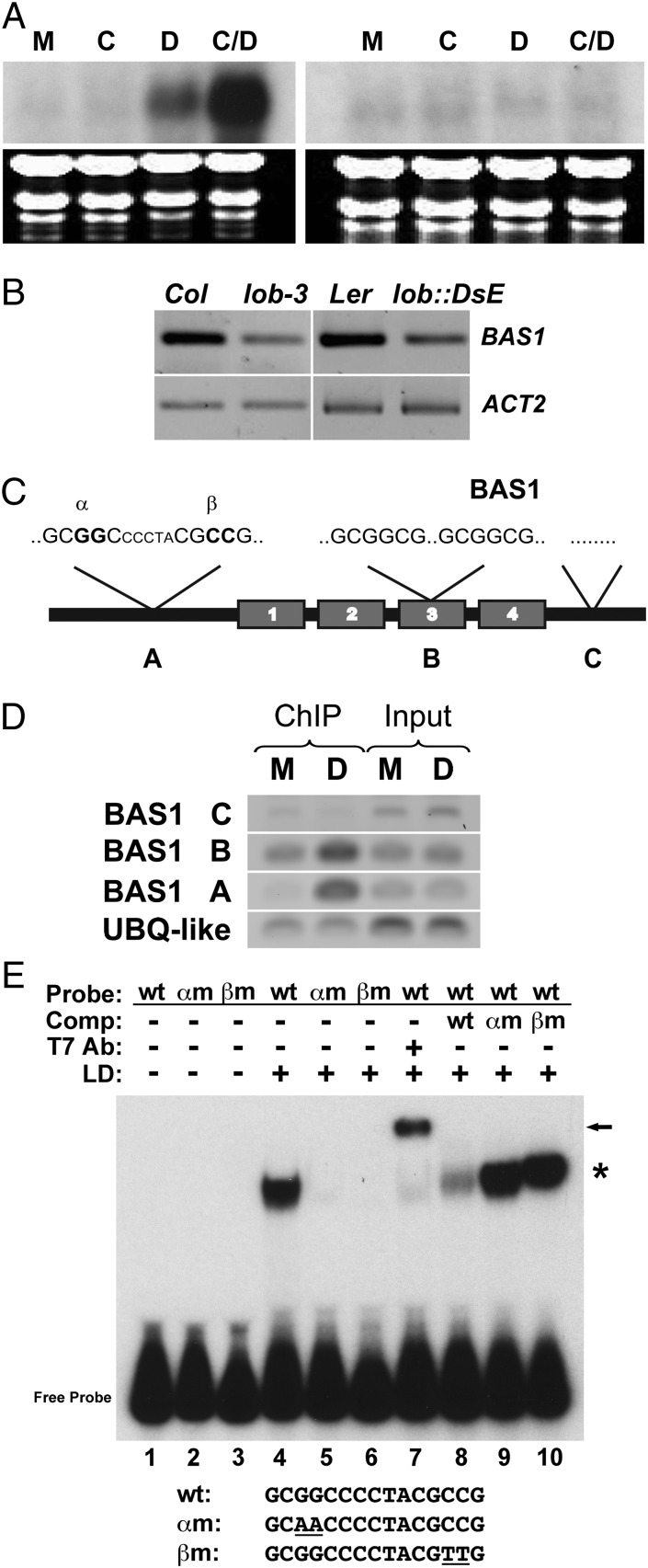
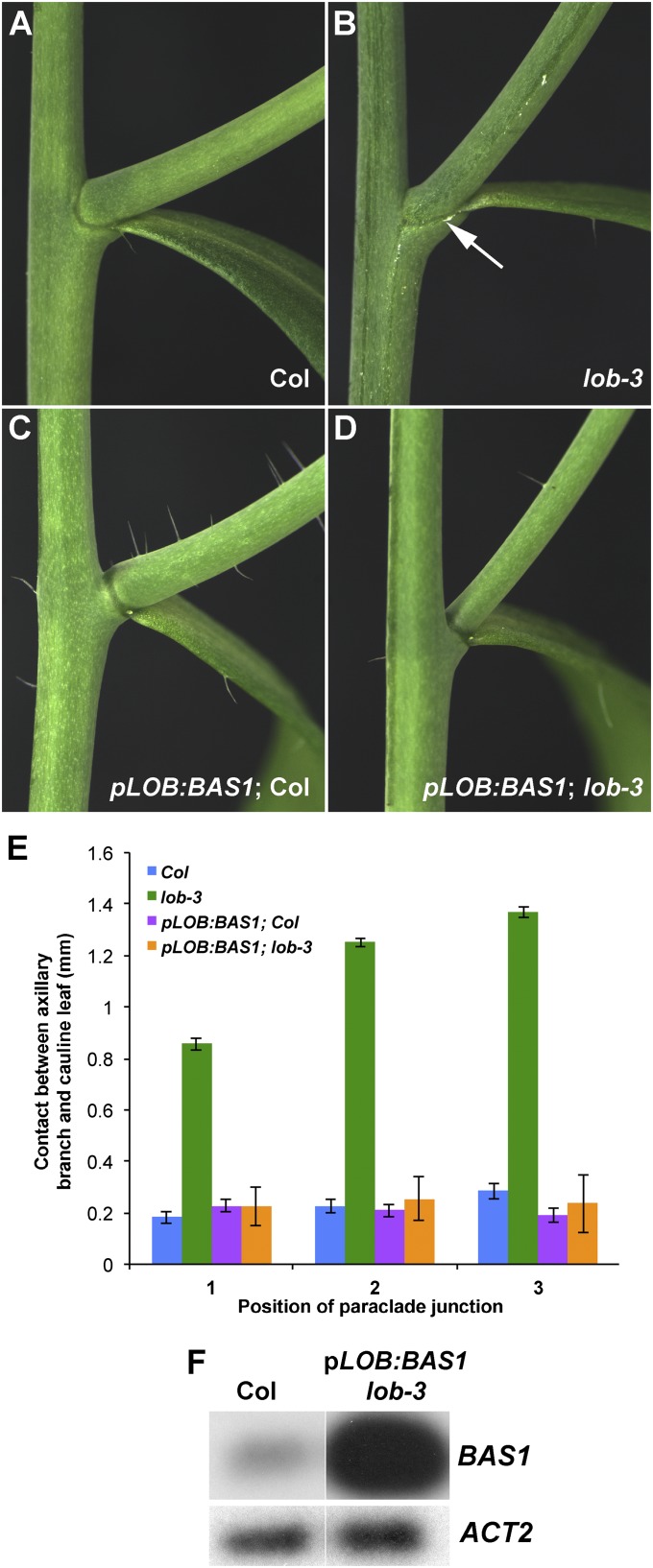
Leaves and flowers begin life as outgrowths from the edges of shoot apical meristems. Stem cell divisions in the meristem center replenish cells that are incorporated into organ primordia at the meristem periphery and leave the meristem. Organ boundaries, regions of limited growth that separate forming organs from the meristem, serve to isolate these two domains and are critical for coordination of organogenesis and meristem maintenance. Boundary formation and maintenance are poorly understood processes, despite the identification of a number of boundary-specific transcription factors. Here we provide genetic and biochemical evidence that the Arabidopsis thaliana transcription factor lateral organ boundaries (LOB) negatively regulates accumulation of the plant steroid hormone brassinosteroid (BR) in organ boundaries. We found that ectopic expression of LOB results in reduced BR responses. We identified BAS1, which encodes a BR-inactivating enzyme, as a direct target of LOB transcriptional activation. Loss-of-function lob mutants exhibit organ fusions, and this phenotype is suppressed by expression of BAS1 under the LOB promoter, indicating that BR hyperaccumulation contributes to the lob mutant phenotype. In addition, LOB expression is BR regulated; therefore, LOB and BR form a feedback loop to modulate local BR accumulation in organ boundaries to limit growth in the boundary domain.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Spatial control of plant steroid signaling.Trends Plant Sci. 2013 May;18(5):235-6. doi: 10.1016/j.tplants.2013.01.005. Epub 2013 Feb 4. Trends Plant Sci. 2013. PMID: 23384602
-
Plant development: brassinosteroids go out of bounds.Curr Biol. 2013 Feb 18;23(4):R152-4. doi: 10.1016/j.cub.2013.01.001. Curr Biol. 2013. PMID: 23428325
References
-
- Barton MK. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev Biol. 2010;341(1):95–113. - PubMed
-
- Hussey G. Cell division and expansion and resultant tissue tensions in shoot apex during formation of a leaf primordium in tomato. J Exp Bot. 1971;22:702–714.
-
- Breuil-Broyer S, et al. High-resolution boundary analysis during Arabidopsis thaliana flower development. Plant J. 2004;38(1):182–192. - PubMed
-
- Rast MI, Simon R. The meristem-to-organ boundary: More than an extremity of anything. Curr Opin Genet Dev. 2008;18(4):287–294. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
