

Topological specificity and hierarchical network of the circadian calcium rhythm in the suprachiasmatic nucleus
- PMID: 23213253
- PMCID: PMC3535646
- DOI: 10.1073/pnas.1214415110
Topological specificity and hierarchical network of the circadian calcium rhythm in the suprachiasmatic nucleus
Abstract
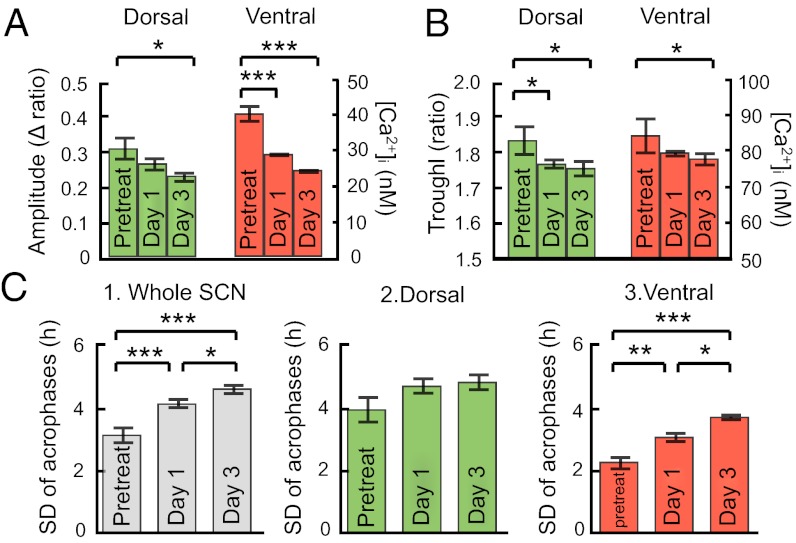
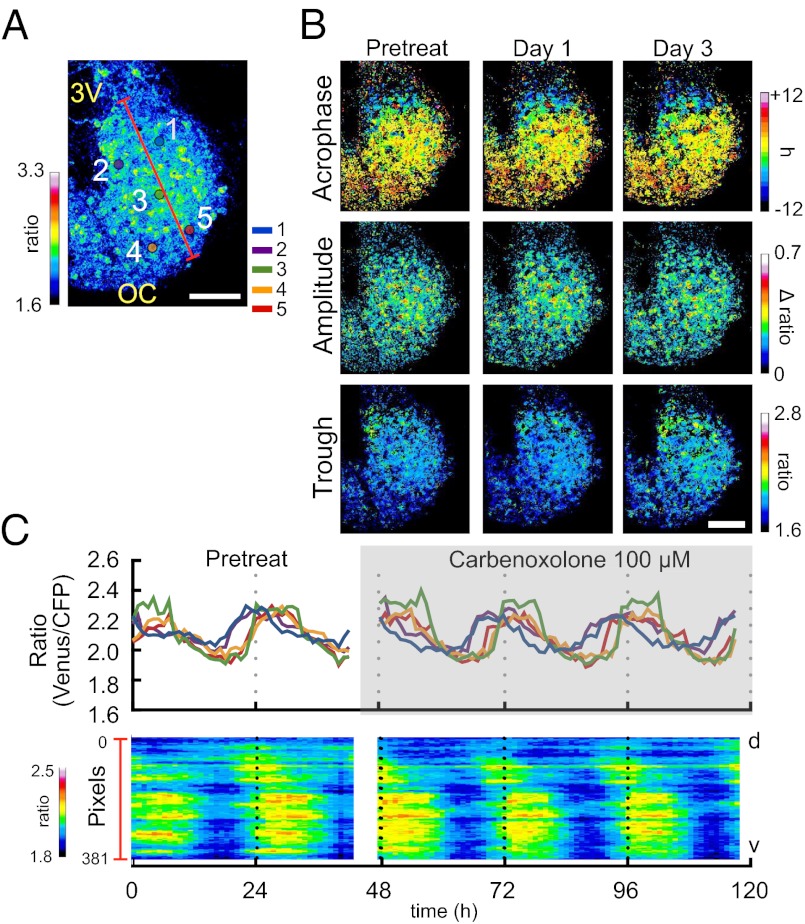
The circadian pacemaker in the hypothalamic suprachiasmatic nucleus (SCN) is a hierarchical multioscillator system in which neuronal networks play crucial roles in expressing coherent rhythms in physiology and behavior. However, our understanding of the neuronal network is still incomplete. Intracellular calcium mediates the input signals, such as phase-resetting stimuli, to the core molecular loop involving clock genes for circadian rhythm generation and the output signals from the loop to various cellular functions, including changes in neurotransmitter release. Using a unique large-scale calcium imaging method with genetically encoded calcium sensors, we visualized intracellular calcium from the entire surface of SCN slice in culture including the regions where autonomous clock gene expression was undetectable. We found circadian calcium rhythms at a single-cell level in the SCN, which were topologically specific with a larger amplitude and more delayed phase in the ventral region than the dorsal. The robustness of the rhythm was reduced but persisted even after blocking the neuronal firing with tetrodotoxin (TTX). Notably, TTX dissociated the circadian calcium rhythms between the dorsal and ventral SCN. In contrast, a blocker of gap junctions, carbenoxolone, had only a minor effect on the calcium rhythms at both the single-cell and network levels. These results reveal the topological specificity of the circadian calcium rhythm in the SCN and the presence of coupled regional pacemakers in the dorsal and ventral regions. Neuronal firings are not necessary for the persistence of the calcium rhythms but indispensable for the hierarchical organization of rhythmicity in the SCN.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
