

Brassinosteroids regulate organ boundary formation in the shoot apical meristem of Arabidopsis
- PMID: 23213257
- PMCID: PMC3529081
- DOI: 10.1073/pnas.1210799110
Brassinosteroids regulate organ boundary formation in the shoot apical meristem of Arabidopsis
Abstract
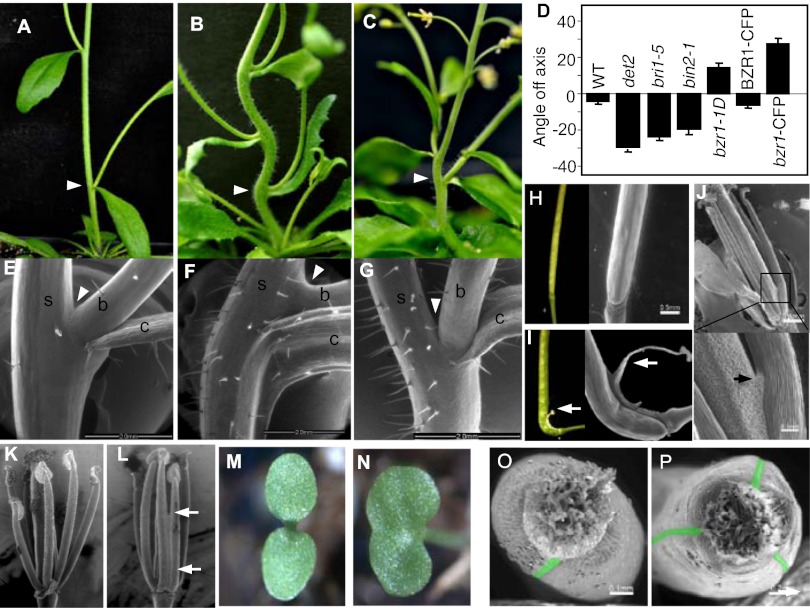
Spatiotemporal control of the formation of organ primordia and organ boundaries from the stem cell niche in the shoot apical meristem (SAM) determines the patterning and architecture of plants, but the underlying signaling mechanisms remain poorly understood. Here we show that brassinosteroids (BRs) play a key role in organ boundary formation by repressing organ boundary identity genes. BR-hypersensitive mutants display organ-fusion phenotypes, whereas BR-insensitive mutants show enhanced organ boundaries. The BR-activated transcription factor BZR1 directly represses the cup-shaped cotyledon (CUC) family of organ boundary identity genes. In WT plants, BZR1 accumulates at high levels in the nuclei of central meristem and organ primordia but at a low level in organ boundary cells to allow CUC gene expression. Activation of BR signaling represses CUC gene expression and causes organ fusion phenotypes. This study uncovers a role for BR in the spatiotemporal control of organ boundary formation and morphogenesis in the SAM.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Comment in
-
Spatial control of plant steroid signaling.Trends Plant Sci. 2013 May;18(5):235-6. doi: 10.1016/j.tplants.2013.01.005. Epub 2013 Feb 4. Trends Plant Sci. 2013. PMID: 23384602
-
Plant development: brassinosteroids go out of bounds.Curr Biol. 2013 Feb 18;23(4):R152-4. doi: 10.1016/j.cub.2013.01.001. Curr Biol. 2013. PMID: 23428325
References
-
- Sablowski R. Plant and animal stem cells: conceptually similar, molecularly distinct? Trends Cell Biol. 2004;14(11):605–611. - PubMed
-
- Sablowski R. The dynamic plant stem cell niches. Curr Opin Plant Biol. 2007;10(6):639–644. - PubMed
-
- Aida M, Tasaka M. Genetic control of shoot organ boundaries. Curr Opin Plant Biol. 2006;9(1):72–77. - PubMed
-
- Lee DK, Geisler M, Springer PS. LATERAL ORGAN FUSION1 and LATERAL ORGAN FUSION2 function in lateral organ separation and axillary meristem formation in Arabidopsis. Development. 2009;136(14):2423–2432. - PubMed
-
- Aida M, Tasaka M. Morphogenesis and patterning at the organ boundaries in the higher plant shoot apex. Plant Mol Biol. 2006;60(6):915–928. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
