

Positive intergenic feedback circuitry, involving EBF1 and FOXO1, orchestrates B-cell fate
- PMID: 23213261
- PMCID: PMC3529039
- DOI: 10.1073/pnas.1211427109
Positive intergenic feedback circuitry, involving EBF1 and FOXO1, orchestrates B-cell fate
Abstract
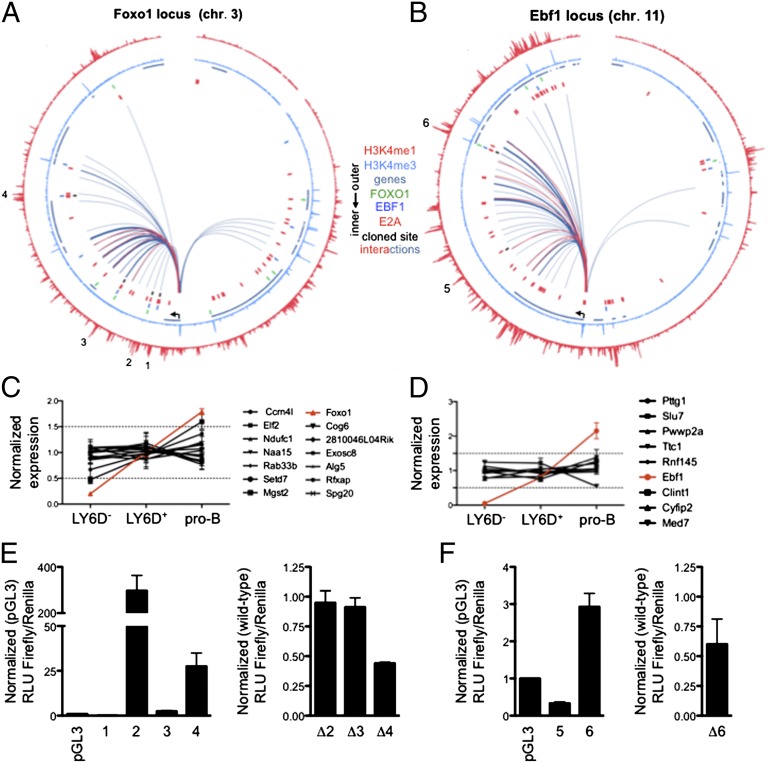
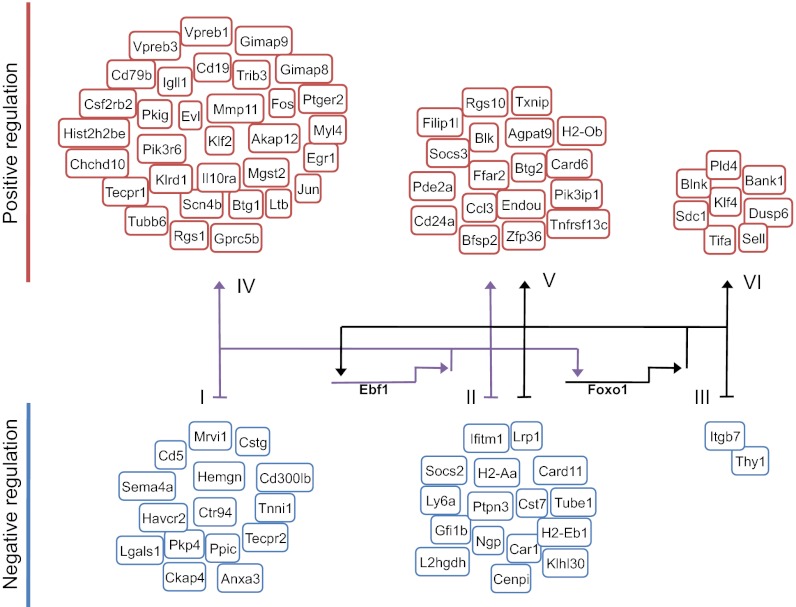
Recent studies have identified a number of transcriptional regulators, including E2A, early B-cell factor 1 (EBF1), FOXO1, and paired box gene 5 (PAX5), that promote early B-cell development. However, how this ensemble of regulators mechanistically promotes B-cell fate remains poorly understood. Here we demonstrate that B-cell development in FOXO1-deficient mice is arrested in the common lymphoid progenitor (CLP) LY6D(+) cell stage. We demonstrate that this phenotype closely resembles the arrest in B-cell development observed in EBF1-deficient mice. Consistent with these observations, we find that the transcription signatures of FOXO1- and EBF1-deficient LY6D(+) progenitors are strikingly similar, indicating a common set of target genes. Furthermore, we found that depletion of EBF1 expression in LY6D(+) CLPs severely affects FOXO1 mRNA abundance, whereas depletion of FOXO1 activity in LY6D(+) CLPs ablates EBF1 transcript levels. We generated a global regulatory network from EBF1 and FOXO1 genome-wide transcription factor occupancy and transcription signatures derived from EBF1- and FOXO1-deficient CLPs. This analysis reveals that EBF1 and FOXO1 act in a positive feedback circuitry to promote and stabilize specification to the B-cell lineage.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Adolfsson J, et al. Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential: A revised road map for adult blood lineage commitment. Cell. 2005;121(2):295–306. - PubMed
-
- Kondo M, Weissman IL, Akashi K. Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell. 1997;91(5):661–672. - PubMed
-
- Mansson R, et al. Single-cell analysis of the common lymphoid progenitor compartment reveals functional and molecular heterogeneity. Blood. 2010;115(13):2601–2609. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
