

Dual pathway spindle assembly increases both the speed and the fidelity of mitosis
- PMID: 23213363
- PMCID: PMC3507165
- DOI: 10.1242/bio.2011012
Dual pathway spindle assembly increases both the speed and the fidelity of mitosis
Abstract
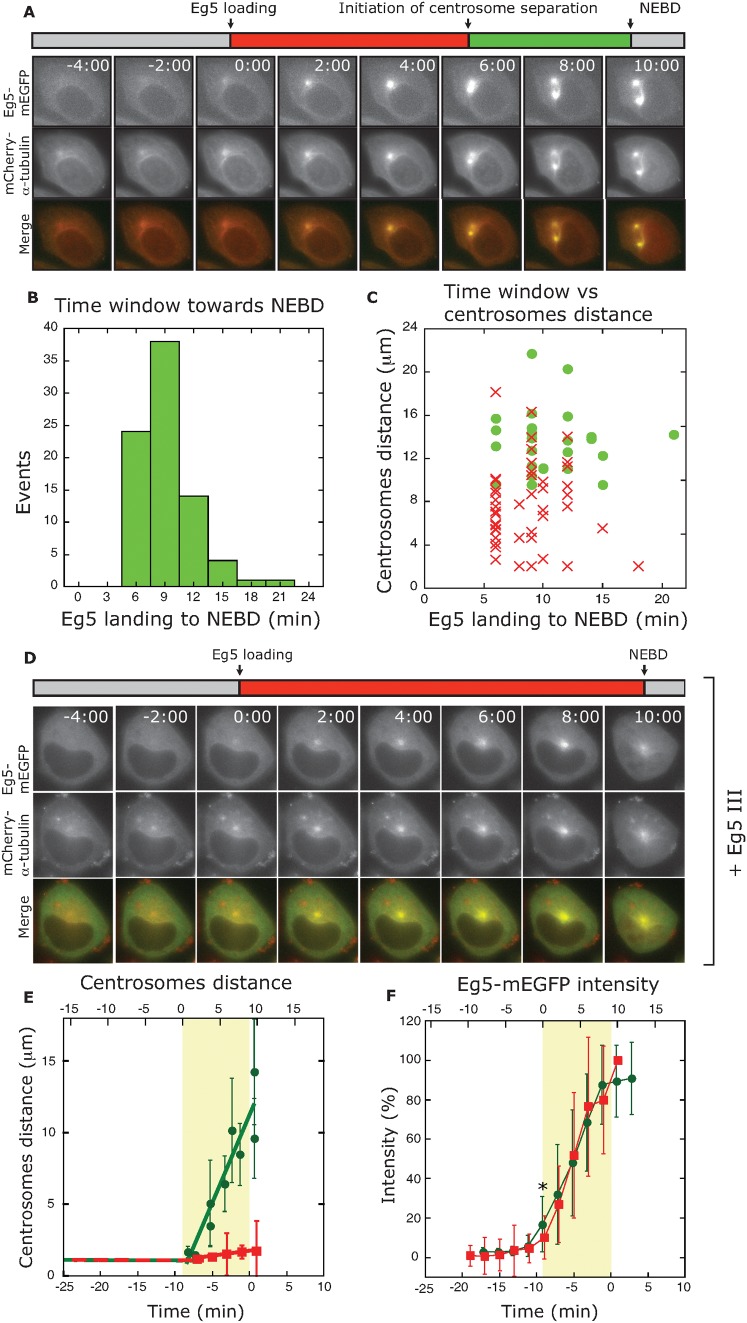
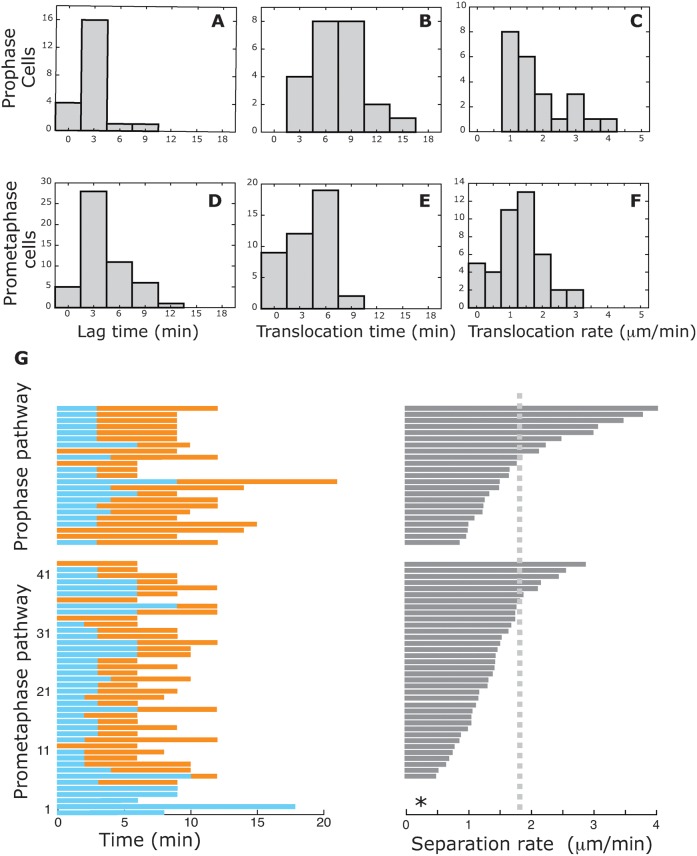
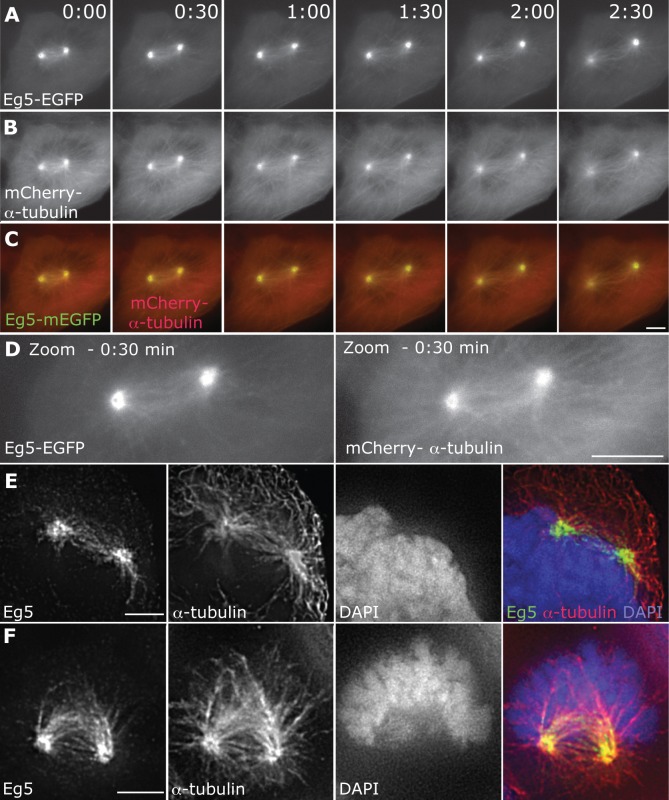
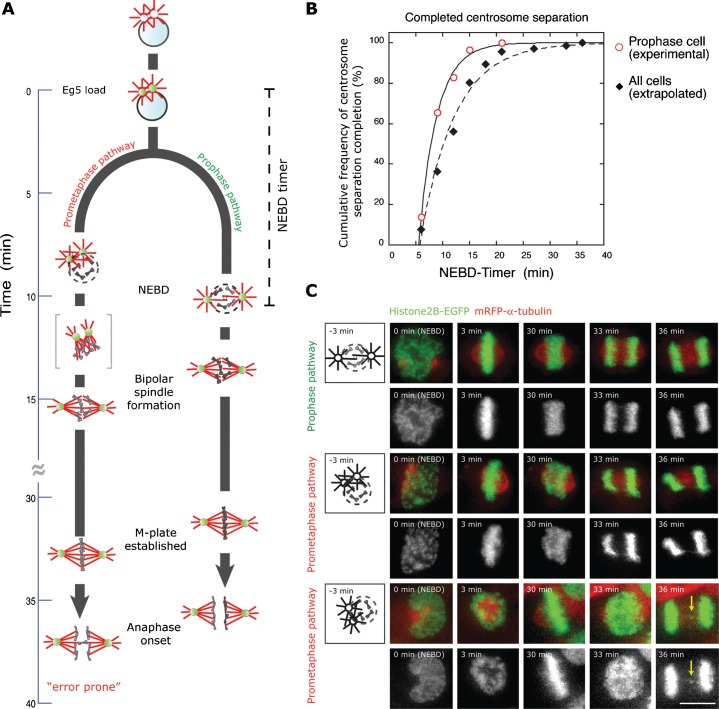
Roughly half of all animal somatic cell spindles assemble by the classical prophase pathway, in which the centrosomes separate ahead of nuclear envelope breakdown (NEBD). The remainder assemble by the prometaphase pathway, in which the centrosomes separate following NEBD. Why cells use dual pathway spindle assembly is unclear. Here, by examining the timing of NEBD relative to the onset of Eg5-mEGFP loading to centrosomes, we show that a time window of 9.2 ± 2.9 min is available for Eg5-driven prophase centrosome separation ahead of NEBD, and that those cells that succeed in separating their centrosomes within this window subsequently show >3-fold fewer chromosome segregation errors and a somewhat faster mitosis. A longer time window would allow more cells to complete prophase centrosome separation and further reduce segregation errors, but at the expense of a slower mitosis. Our data reveal dual pathway mitosis in a new light, as a substantive strategy that increases both the speed and the fidelity of mitosis.
Conflict of interest statement
Figures
References
LinkOut - more resources
Full Text Sources
