

The Aplnr GPCR regulates myocardial progenitor development via a novel cell-non-autonomous, Gα(i/o) protein-independent pathway
- PMID: 23213418
- PMCID: PMC3507289
- DOI: 10.1242/bio.2012380
The Aplnr GPCR regulates myocardial progenitor development via a novel cell-non-autonomous, Gα(i/o) protein-independent pathway
Abstract
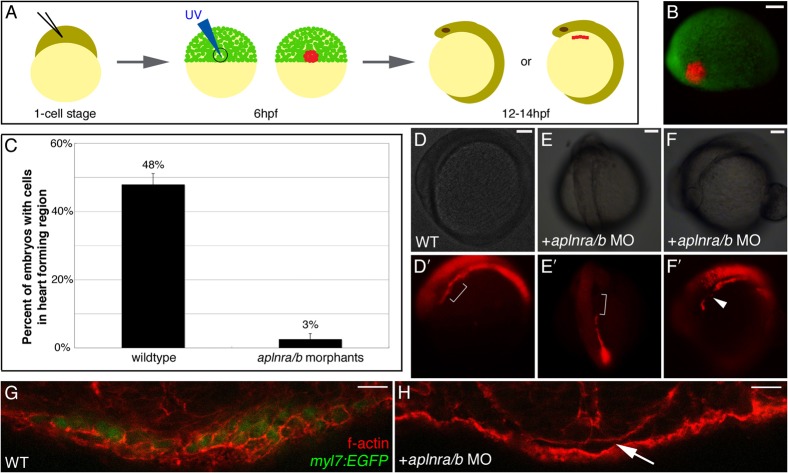
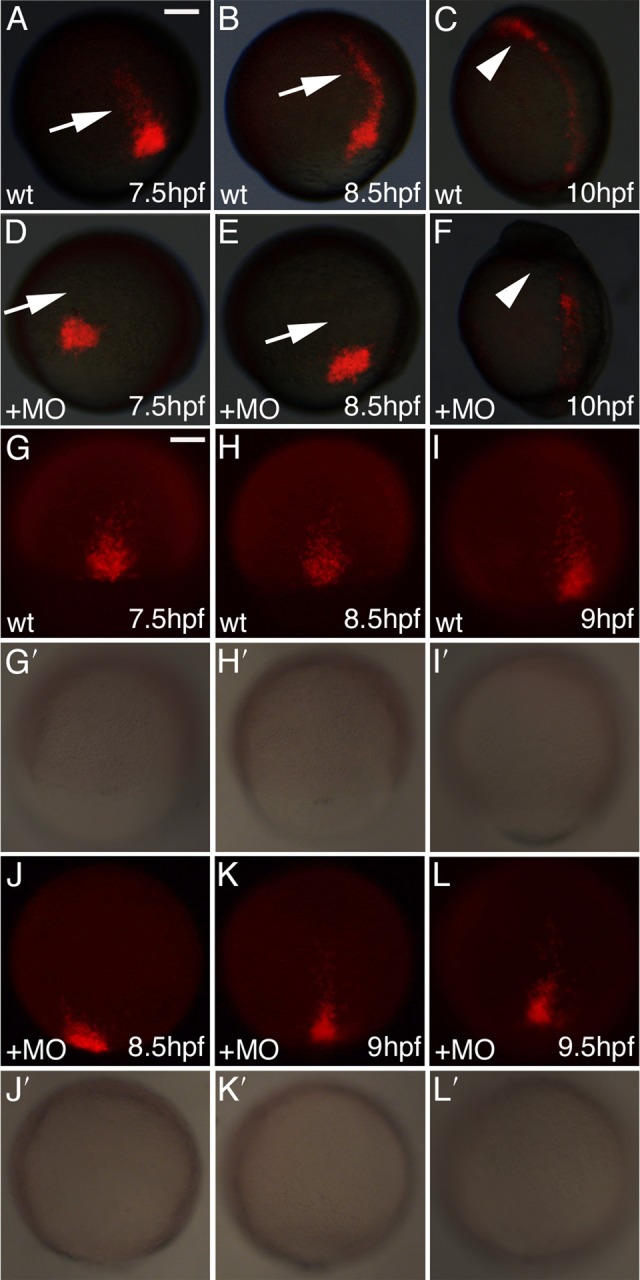
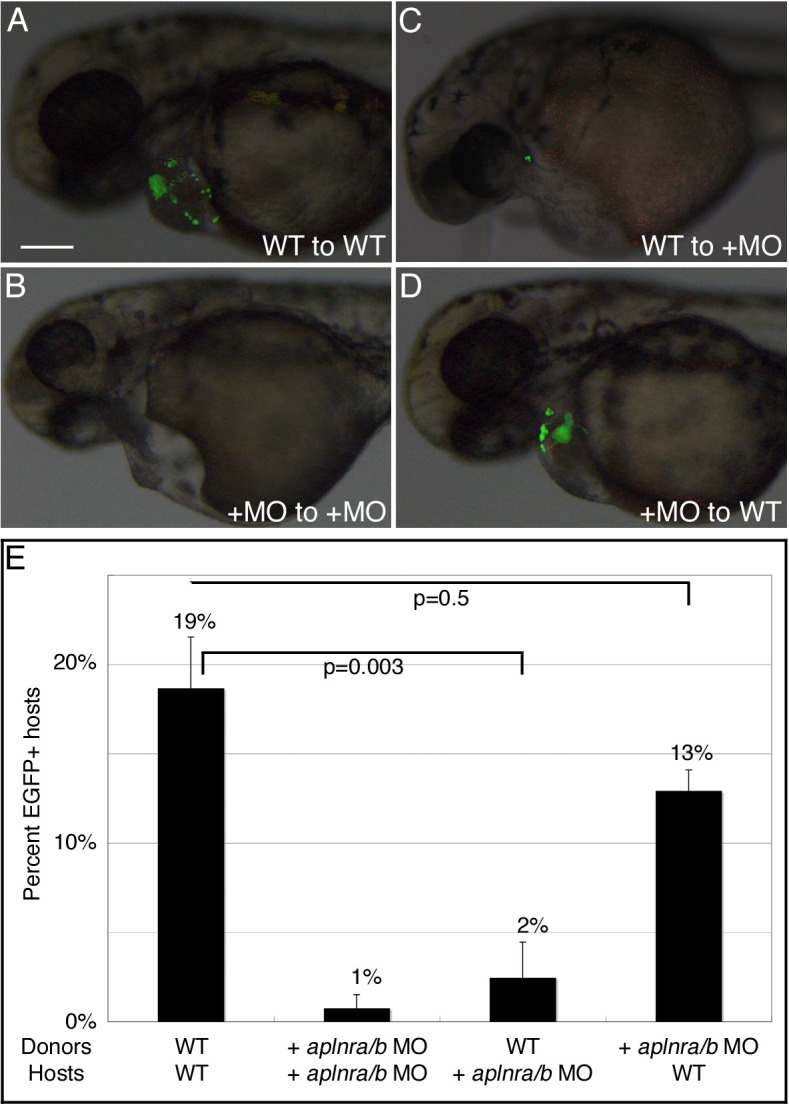
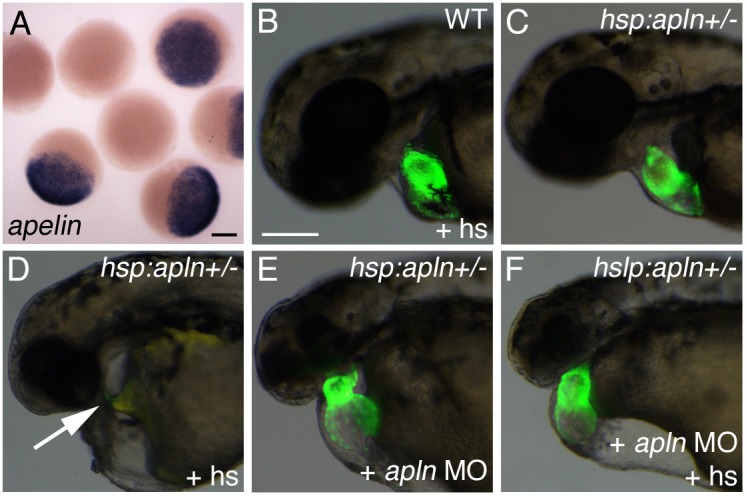
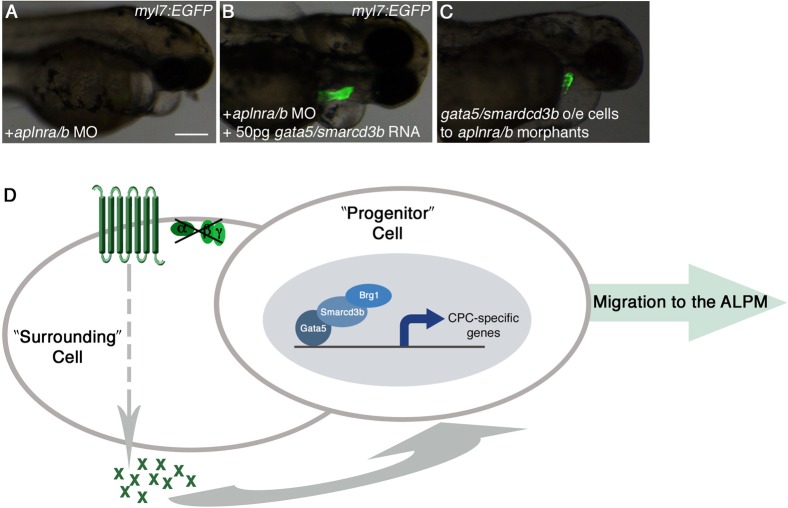
Myocardial progenitor development involves the migration of cells to the anterior lateral plate mesoderm (ALPM) where they are exposed to the necessary signals for heart development to proceed. Whether the arrival of cells to this location is sufficient, or whether earlier signaling events are required, for progenitor development is poorly understood. Here we demonstrate that in the absence of Aplnr signaling, cells fail to migrate to the heart-forming region of the ALPM. Our work uncovers a previously uncharacterized cell-non-autonomous function for Aplnr signaling in cardiac development. Furthermore, we show that both the single known Aplnr ligand, Apelin, and the canonical Gα(i/o) proteins that signal downstream of Aplnr are dispensable for Aplnr function in the context of myocardial progenitor development. This novel Aplnr signal can be substituted for by activation of Gata5/Smarcd3 in myocardial progenitors, suggesting a novel mechanism for Aplnr signaling in the establishment of a niche required for the proper migration/development of myocardial progenitor cells.
Keywords: Apelin Receptor; Gastrulation; Myocardial Progenitors; Zebrafish.
Figures

Similar articles
-
The Apelin receptor enhances Nodal/TGFβ signaling to ensure proper cardiac development.Elife. 2016 Apr 14;5:e13758. doi: 10.7554/eLife.13758. Elife. 2016. PMID: 27077952 Free PMC article.
-
Apelin Receptor Signaling During Mesoderm Development.Adv Exp Med Biol. 2020;1298:1-15. doi: 10.1007/5584_2020_567. Adv Exp Med Biol. 2020. PMID: 32648246
-
Apelin receptor (Aplnr) signaling promotes fibroblast migration.Tissue Cell. 2019 Feb;56:98-106. doi: 10.1016/j.tice.2019.01.003. Epub 2019 Jan 17. Tissue Cell. 2019. PMID: 30736911
-
The Apelin/APLNR system modulates tumor immune response by reshaping the tumor microenvironment.Gene. 2022 Aug 5;834:146564. doi: 10.1016/j.gene.2022.146564. Epub 2022 May 20. Gene. 2022. PMID: 35598689 Review.
-
APLN/APLNR Signaling Controls Key Pathological Parameters of Glioblastoma.Cancers (Basel). 2021 Aug 2;13(15):3899. doi: 10.3390/cancers13153899. Cancers (Basel). 2021. PMID: 34359800 Free PMC article. Review.
Cited by
-
Nodal signaling has dual roles in fate specification and directed migration during germ layer segregation in zebrafish.Development. 2018 Sep 14;145(17):dev163535. doi: 10.1242/dev.163535. Development. 2018. PMID: 30111654 Free PMC article.
-
Aplnra/b Sequentially Regulate Organ Left-Right Patterning via Distinct Mechanisms.Int J Biol Sci. 2019 May 11;15(6):1225-1239. doi: 10.7150/ijbs.30100. eCollection 2019. Int J Biol Sci. 2019. PMID: 31223282 Free PMC article.
-
A self-generated Toddler gradient guides mesodermal cell migration.Sci Adv. 2022 Sep 16;8(37):eadd2488. doi: 10.1126/sciadv.add2488. Epub 2022 Sep 14. Sci Adv. 2022. PMID: 36103529 Free PMC article.
-
The Apelinergic System in Pregnancy.Int J Mol Sci. 2023 Apr 28;24(9):8014. doi: 10.3390/ijms24098014. Int J Mol Sci. 2023. PMID: 37175743 Free PMC article. Review.
-
Cellular and molecular mechanisms of convergence and extension in zebrafish.Curr Top Dev Biol. 2020;136:377-407. doi: 10.1016/bs.ctdb.2019.08.001. Epub 2019 Sep 3. Curr Top Dev Biol. 2020. PMID: 31959296 Free PMC article. Review.
References
-
- Ashley E. A., Powers J., Chen M., Kundu R., Finsterbach T., Caffarelli A., Deng A., Eichhorn J., Mahajan R., Agrawal R. et al. (2005). The endogenous peptide apelin potently improves cardiac contractility and reduces cardiac loading in vivo. Cardiovasc. Res. 65, 73–82 10.1016/j.cardiores.2004.08.018 - DOI - PMC - PubMed
-
- Berghmans S., Murphey R. D., Wienholds E., Neuberg D., Kutok J. L., Fletcher C. D., Morris J. P., Liu T. X., Schulte-Merker S., Kanki J. P. et al. (2005). tp53 mutant zebrafish develop malignant peripheral nerve sheath tumors. Proc. Natl. Acad. Sci. USA 102, 407–412 10.1073/pnas.0406252102 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
