

Rearrangements between differentiating hair cells coordinate planar polarity and the establishment of mirror symmetry in lateral-line neuromasts
- PMID: 23213442
- PMCID: PMC3507212
- DOI: 10.1242/bio.2012570
Rearrangements between differentiating hair cells coordinate planar polarity and the establishment of mirror symmetry in lateral-line neuromasts
Abstract
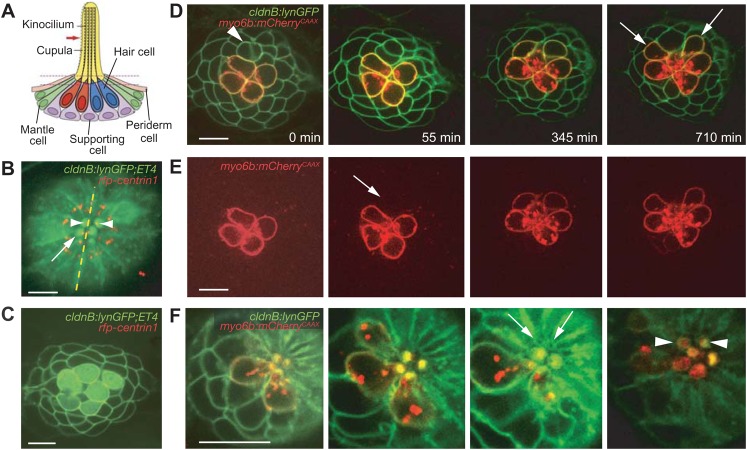
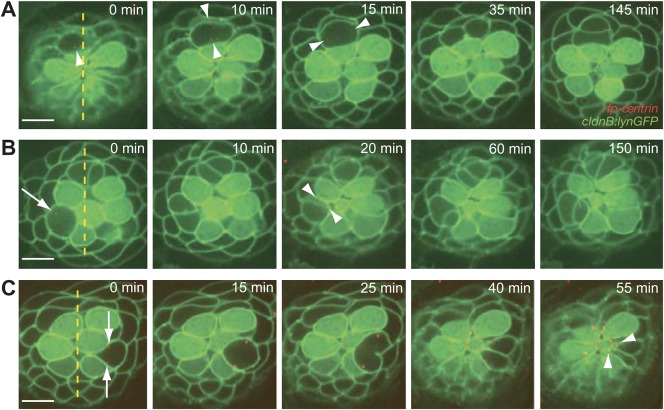
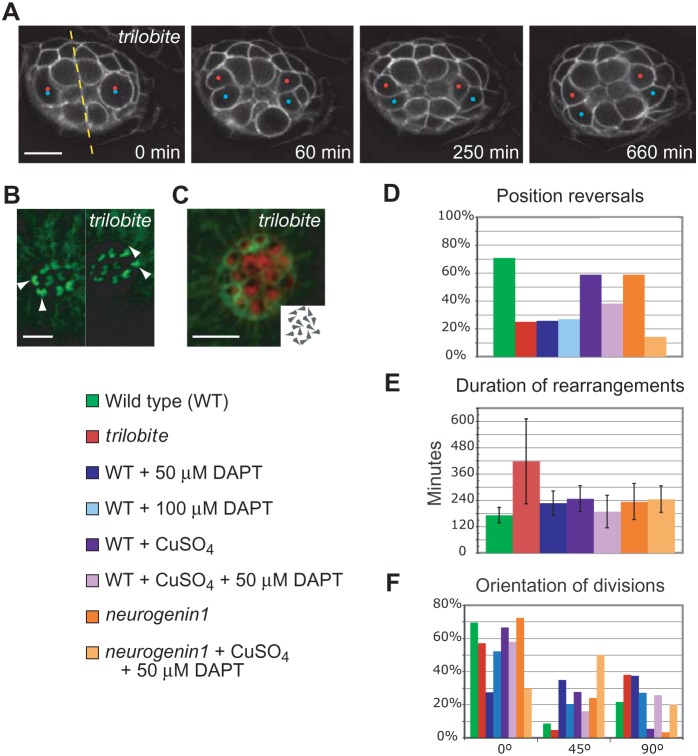
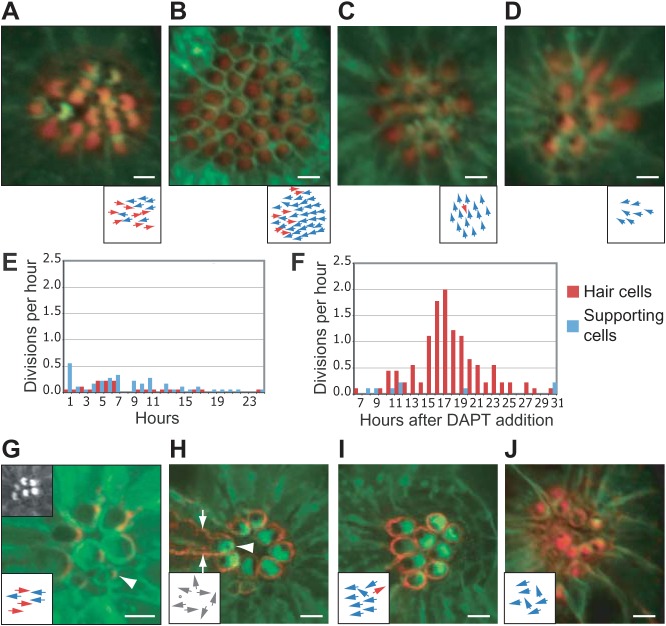
In addition to their ubiquitous apical-basal polarity, many epithelia are also polarized along an orthogonal axis, a phenomenon termed planar cell polarity (PCP). In the mammalian inner ear and the zebrafish lateral line, PCP is revealed through the orientation of mechanosensitive hair cells relative to each other and to the body axes. In each neuromast, the receptor organ of the lateral line, hair bundles are arranged in a mirror-symmetrical fashion. Here we show that the establishment of mirror symmetry is preceded by rotational rearrangements between hair-cell pairs, a behavior consistently associated with the division of hair-cell precursors. Time-lapse imaging of trilobite mutants, which lack the core PCP constituent Vang-like protein 2 (Vangl2), shows that their misoriented hair cells correlate with misaligned divisions of hair-cell precursors and an inability to complete rearrangements accurately. Vangl2 is asymmetrically localized in the cells of the neuromast, a configuration required for accurate completion of rearrangements. Manipulation of Vangl2 expression or of Notch signaling results in a uniform hair-cell polarity, indicating that rearrangements refine neuromast polarity with respect to the body axes.
Keywords: Neurogenin; Notch; Planar cell polarity; Vang-like protein 2.
Conflict of interest statement
Figures
References
-
- Blader P., Fischer N., Gradwohl G., Guillemot F., Strähle U. (1997). The activity of neurogenin1 is controlled by local cues in the zebrafish embryo. Development 124, 4557–4569. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
