

Life-long preservation of the regenerative capacity in the fin and heart in zebrafish
- PMID: 23213467
- PMCID: PMC3507221
- DOI: 10.1242/bio.20121057
Life-long preservation of the regenerative capacity in the fin and heart in zebrafish
Abstract
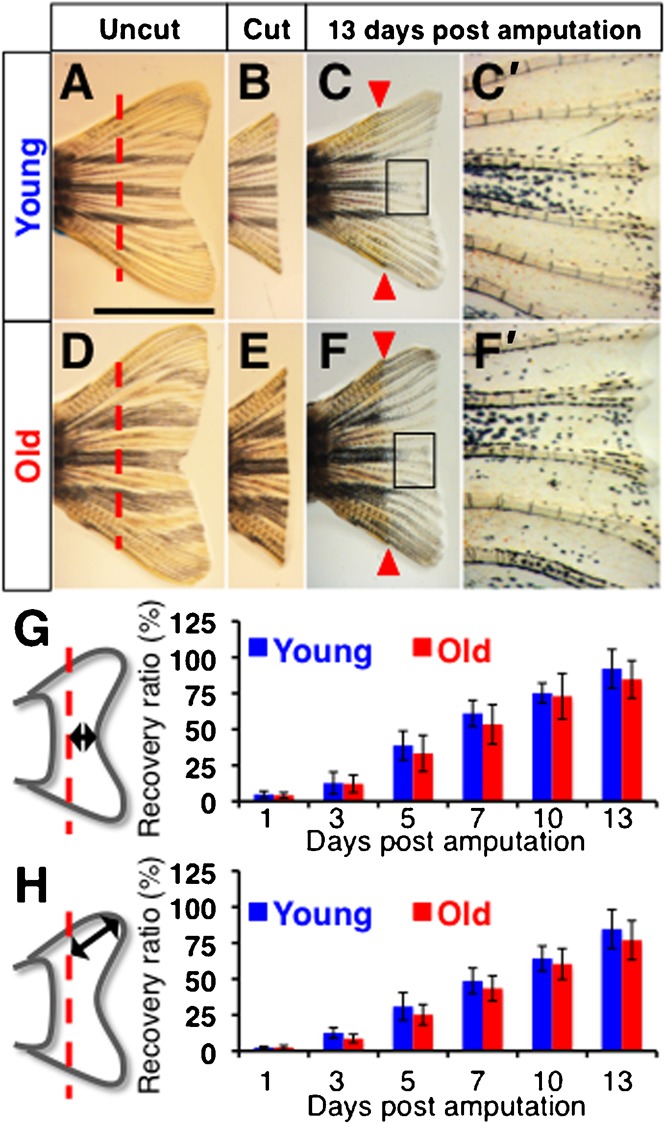
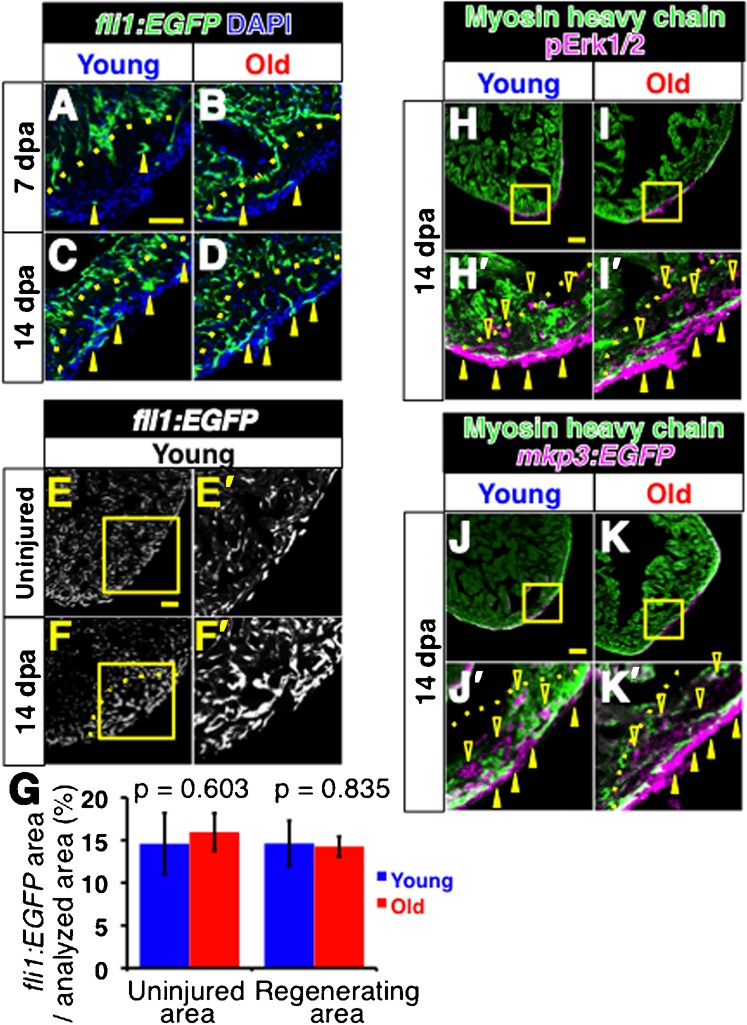
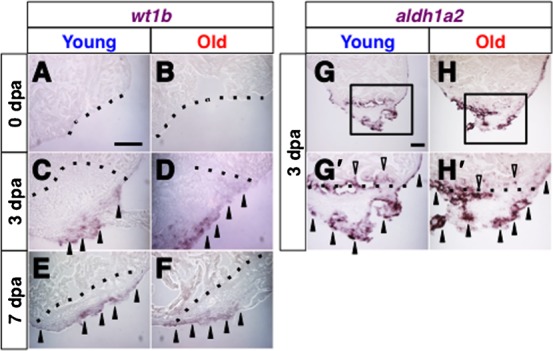
The zebrafish is a widely used model animal to study the regeneration of organs, such as the fin and heart. Their average lifetime is about 3 years, and recent studies have shown that zebrafish exhibit aging-related degeneration, suggesting the possibility that aging might affect regenerative potential. In order to investigate this possibility, we compared regeneration of the fin and heart after experimental amputation in young (6-12 month old) and old (26-36 month old) fish. Comparison of recovery rate of the caudal fin, measured every two or three days from one day post amputation until 13 days post amputation, show that fins in young and old fish regenerate at a similar rate. In the heart, myocardium regeneration and cardiomyocyte proliferation occurred similarly in the two groups. Moreover, neo-vascularization, as well as activation of fibroblast growth factor signaling, which is required for neo-vascularization, occurred similarly. The epicardial tissue is a thin layer tissue that covers the heart, and starts to express several genes immediately in response to injury. The expression of epicardial genes, such as wt1b and aldh1a2, in response to heart injury was comparable in two groups. Our results demonstrate that zebrafish preserve a life-long regenerative ability of the caudal fin and heart.
Keywords: Aging; Fin; Heart; Regeneration; Zebrafish.
Conflict of interest statement
Figures


References
-
- Akimenko M.-A., Johnson S. L., Westerfield M., Ekker M. (1995). Differential induction of four msx homeobox genes during fin development and regeneration in zebrafish. Development 121, 347–357. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
