

Mammalian formin Fhod3 plays an essential role in cardiogenesis by organizing myofibrillogenesis
- PMID: 23213483
- PMCID: PMC3507241
- DOI: 10.1242/bio.20121370
Mammalian formin Fhod3 plays an essential role in cardiogenesis by organizing myofibrillogenesis
Abstract
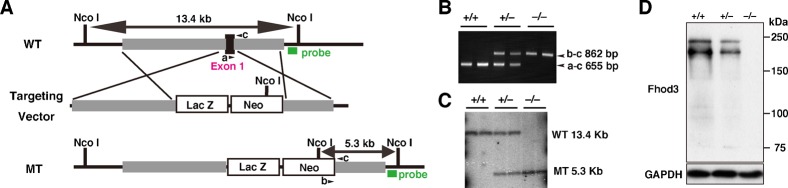
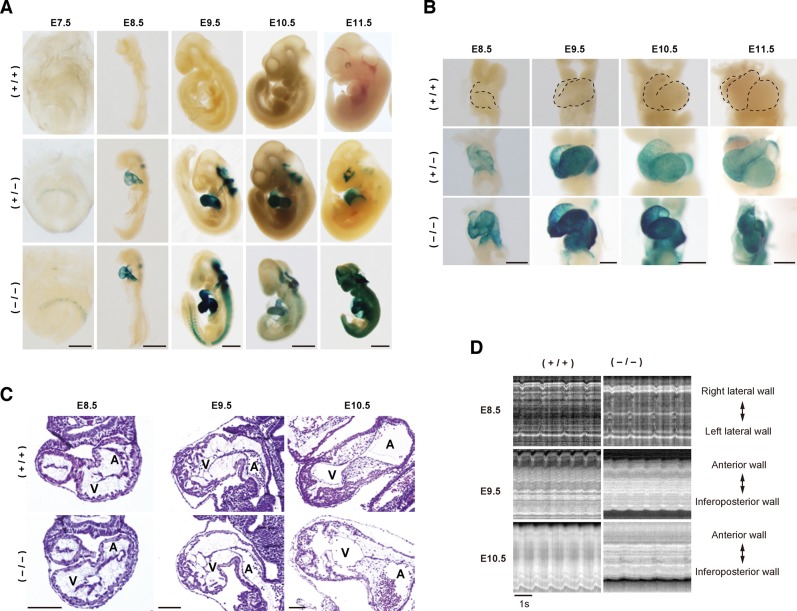
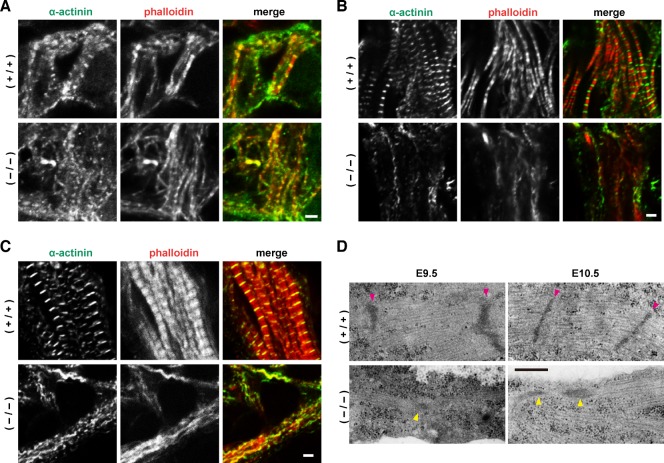
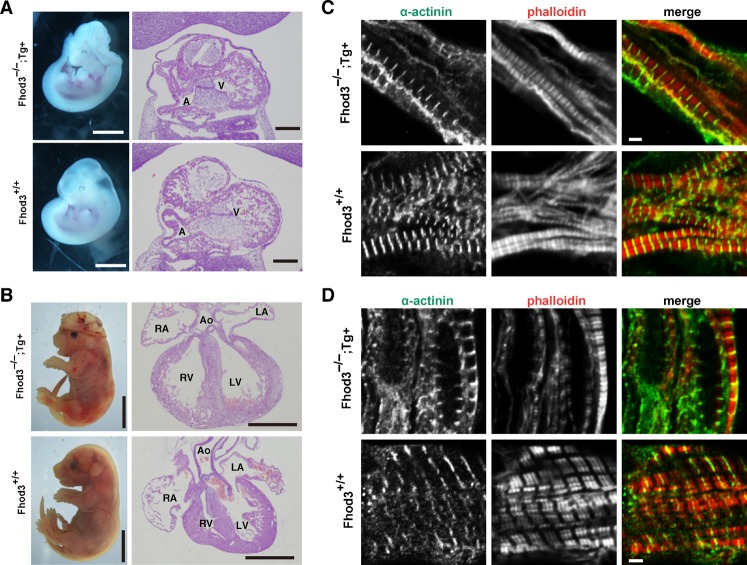
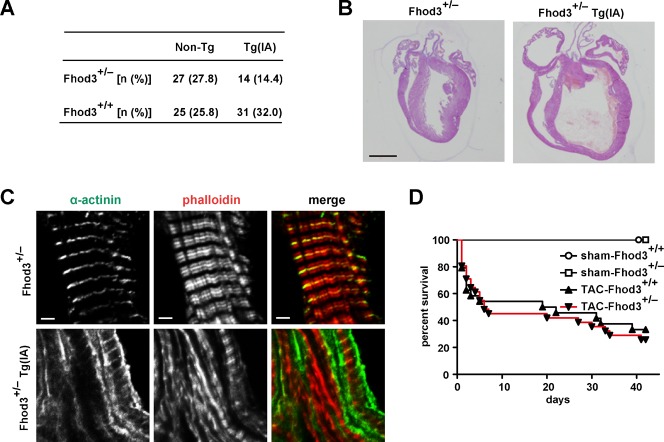
Heart development requires organized integration of actin filaments into the sarcomere, the contractile unit of myofibrils, although it remains largely unknown how actin filaments are assembled during myofibrillogenesis. Here we show that Fhod3, a member of the formin family of proteins that play pivotal roles in actin filament assembly, is essential for myofibrillogenesis at an early stage of heart development. Fhod3(-/-) mice appear normal up to embryonic day (E) 8.5, when the developing heart, composed of premyofibrils, initiates spontaneous contraction. However, these premyofibrils fail to mature and myocardial development does not continue, leading to embryonic lethality by E11.5. Transgenic expression of wild-type Fhod3 in the heart restores myofibril maturation and cardiomyogenesis, which allow Fhod3(-/-) embryos to develop further. Moreover, cardiomyopathic changes with immature myofibrils are caused in mice overexpressing a mutant Fhod3, defective in binding to actin. These findings indicate that actin dynamics, regulated by Fhod3, participate in sarcomere organization during myofibrillogenesis and thus play a crucial role in heart development.
Keywords: Actin; Fhod3; Formin; Myofibrillogenesis; Sarcomere.
Conflict of interest statement
Figures
Similar articles
-
Transgenic Expression of the Formin Protein Fhod3 Selectively in the Embryonic Heart: Role of Actin-Binding Activity of Fhod3 and Its Sarcomeric Localization during Myofibrillogenesis.PLoS One. 2016 Feb 5;11(2):e0148472. doi: 10.1371/journal.pone.0148472. eCollection 2016. PLoS One. 2016. PMID: 26848968 Free PMC article.
-
The actin-organizing formin protein Fhod3 is required for postnatal development and functional maintenance of the adult heart in mice.J Biol Chem. 2018 Jan 5;293(1):148-162. doi: 10.1074/jbc.M117.813931. Epub 2017 Nov 20. J Biol Chem. 2018. PMID: 29158260 Free PMC article.
-
Expression and subcellular localization of mammalian formin Fhod3 in the embryonic and adult heart.PLoS One. 2012;7(4):e34765. doi: 10.1371/journal.pone.0034765. Epub 2012 Apr 11. PLoS One. 2012. PMID: 22509354 Free PMC article.
-
Assembly and Maintenance of Myofibrils in Striated Muscle.Handb Exp Pharmacol. 2017;235:39-75. doi: 10.1007/164_2016_53. Handb Exp Pharmacol. 2017. PMID: 27832381 Review.
-
Actin in striated muscle: recent insights into assembly and maintenance.Biophys Rev. 2012 Mar;4(1):17-25. doi: 10.1007/s12551-011-0062-7. Epub 2011 Dec 20. Biophys Rev. 2012. PMID: 28510000 Free PMC article. Review.
Cited by
-
Actin-Binding Proteins in Cardiac Hypertrophy.Cells. 2022 Nov 11;11(22):3566. doi: 10.3390/cells11223566. Cells. 2022. PMID: 36428995 Free PMC article. Review.
-
Loss of Sarcomere-associated Formins Disrupts Z-line Organization, but does not Prevent Thin Filament Assembly in Caenorhabditis elegans Muscle.J Cytol Histol. 2015 Mar;6(2):318. doi: 10.4172/2157-7099.1000318. J Cytol Histol. 2015. PMID: 26161293 Free PMC article.
-
Transgenic Expression of the Formin Protein Fhod3 Selectively in the Embryonic Heart: Role of Actin-Binding Activity of Fhod3 and Its Sarcomeric Localization during Myofibrillogenesis.PLoS One. 2016 Feb 5;11(2):e0148472. doi: 10.1371/journal.pone.0148472. eCollection 2016. PLoS One. 2016. PMID: 26848968 Free PMC article.
-
FHOD formin and SRF promote post-embryonic striated muscle growth through separate pathways in C. elegans.Exp Cell Res. 2021 Jan 1;398(1):112388. doi: 10.1016/j.yexcr.2020.112388. Epub 2020 Nov 20. Exp Cell Res. 2021. PMID: 33221314 Free PMC article.
-
The expression of the formin Fhod3 in mouse tongue striated muscle.Cell Struct Funct. 2024 Nov 7;49(2):111-122. doi: 10.1247/csf.24044. Epub 2024 Oct 10. Cell Struct Funct. 2024. PMID: 39384365 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
