

Chronic caloric restriction preserves mitochondrial function in senescence without increasing mitochondrial biogenesis
- PMID: 23217257
- PMCID: PMC3544078
- DOI: 10.1016/j.cmet.2012.11.003
Chronic caloric restriction preserves mitochondrial function in senescence without increasing mitochondrial biogenesis
Abstract
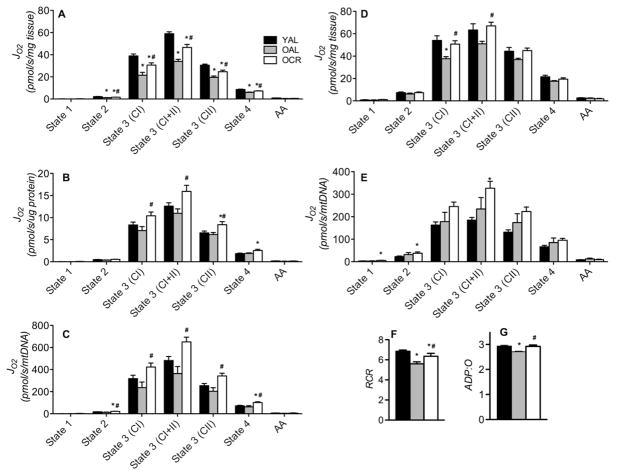
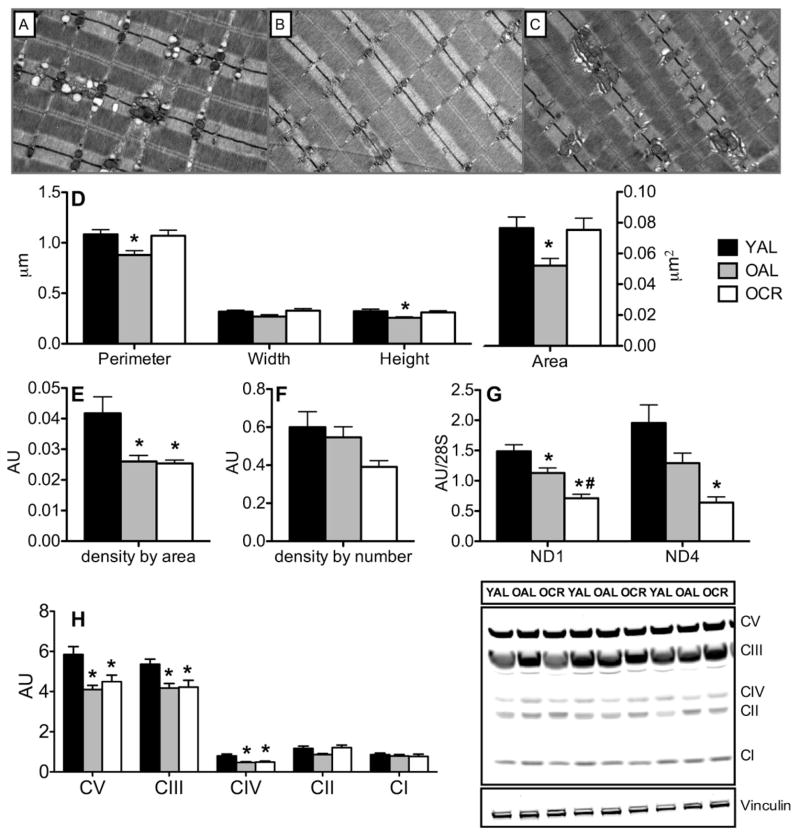
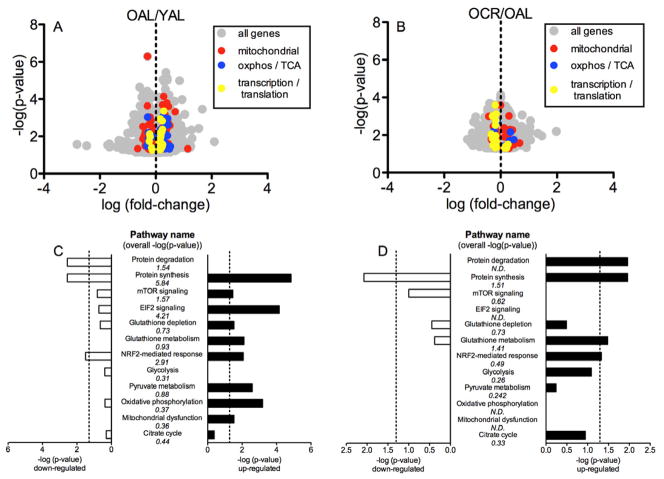
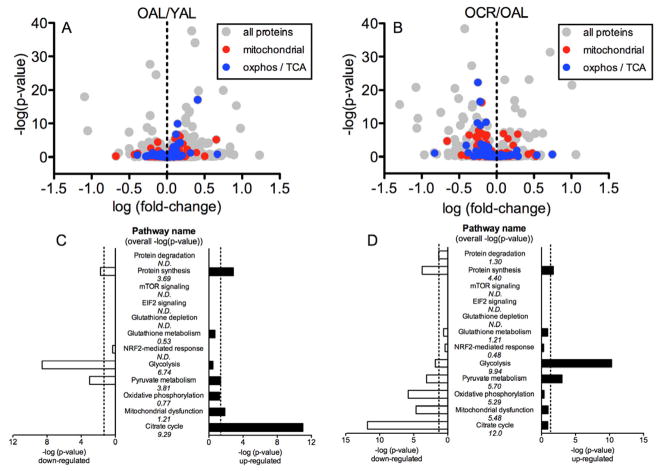
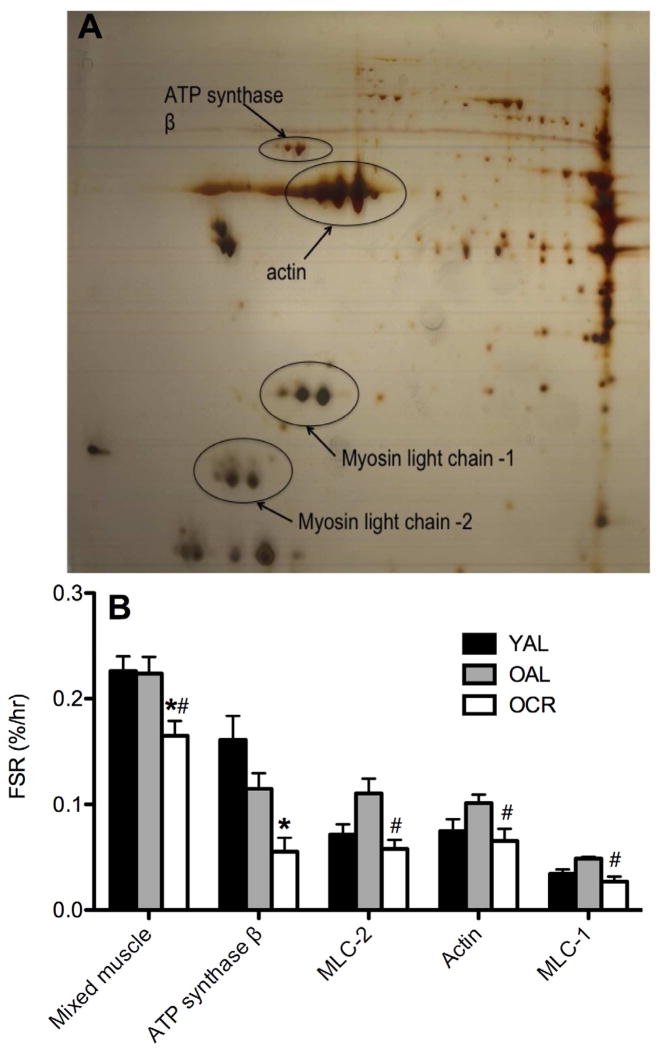
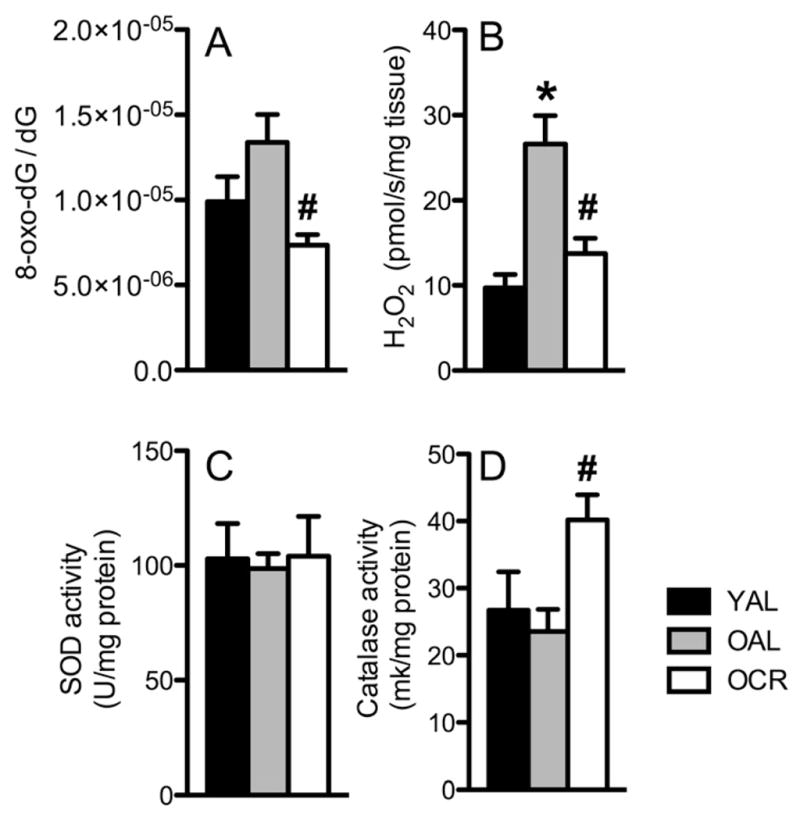
Caloric restriction (CR) mitigates many detrimental effects of aging and prolongs life span. CR has been suggested to increase mitochondrial biogenesis, thereby attenuating age-related declines in mitochondrial function, a concept that is challenged by recent studies. Here we show that lifelong CR in mice prevents age-related loss of mitochondrial oxidative capacity and efficiency, measured in isolated mitochondria and permeabilized muscle fibers. We find that these beneficial effects of CR occur without increasing mitochondrial abundance. Whole-genome expression profiling and large-scale proteomic surveys revealed expression patterns inconsistent with increased mitochondrial biogenesis, which is further supported by lower mitochondrial protein synthesis with CR. We find that CR decreases oxidant emission, increases antioxidant scavenging, and minimizes oxidative damage to DNA and protein. These results demonstrate that CR preserves mitochondrial function by protecting the integrity and function of existing cellular components rather than by increasing mitochondrial biogenesis.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Baker DJ, Betik AC, Krause DJ, Hepple RT. No decline in skeletal muscle oxidative capacity with aging in long-term calorically restricted rats: effects are independent of mitochondrial DNA integrity. J Gerontol A Biol Sci Med Sci. 2006;61:675–684. - PubMed
-
- Cahill GF., Jr Starvation in man. N Engl J Med. 1970;282:668–675. - PubMed
-
- Calleja M, Pena P, Ugalde C, Ferreiro C, Marco R, Garesse R. Mitochondrial DNA remains intact during Drosophila aging, but the levels of mitochondrial transcripts are significantly reduced. J Biol Chem. 1993;268:18891–18897. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
