

Circadian oscillations of protein-coding and regulatory RNAs in a highly dynamic mammalian liver epigenome
- PMID: 23217262
- PMCID: PMC3541940
- DOI: 10.1016/j.cmet.2012.11.004
Circadian oscillations of protein-coding and regulatory RNAs in a highly dynamic mammalian liver epigenome
Abstract
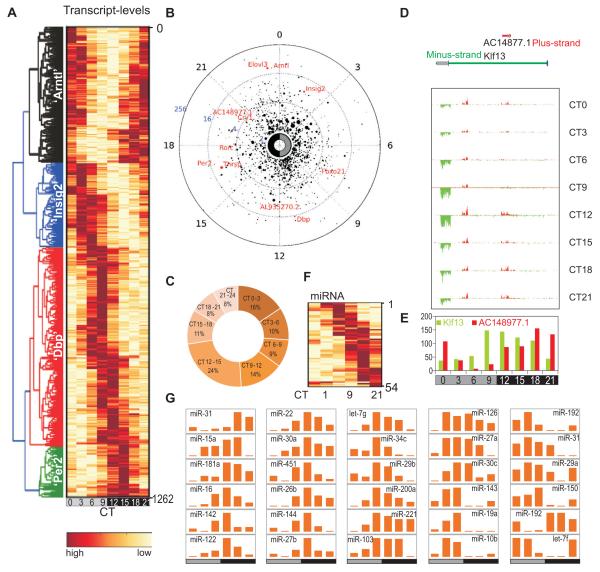
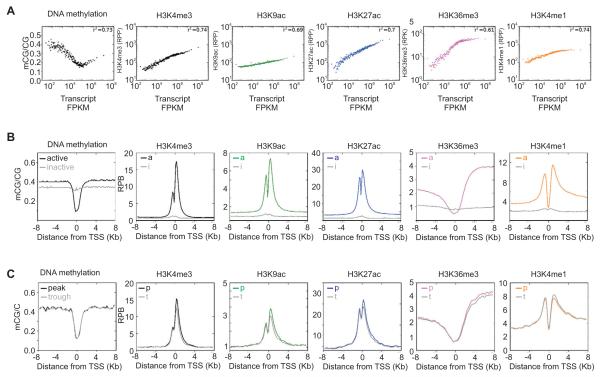
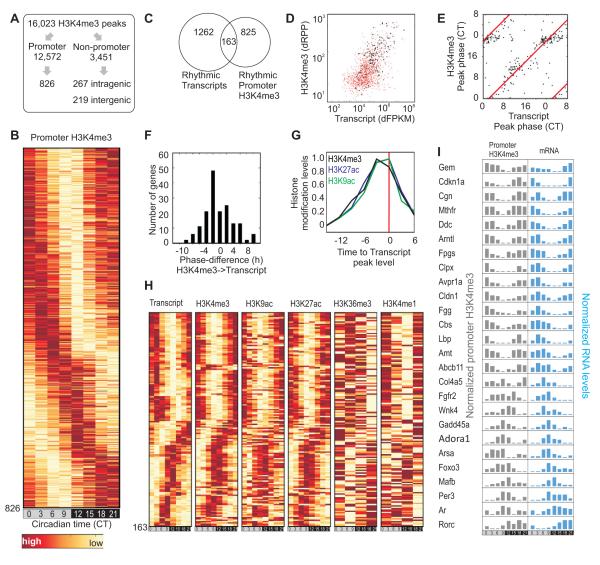
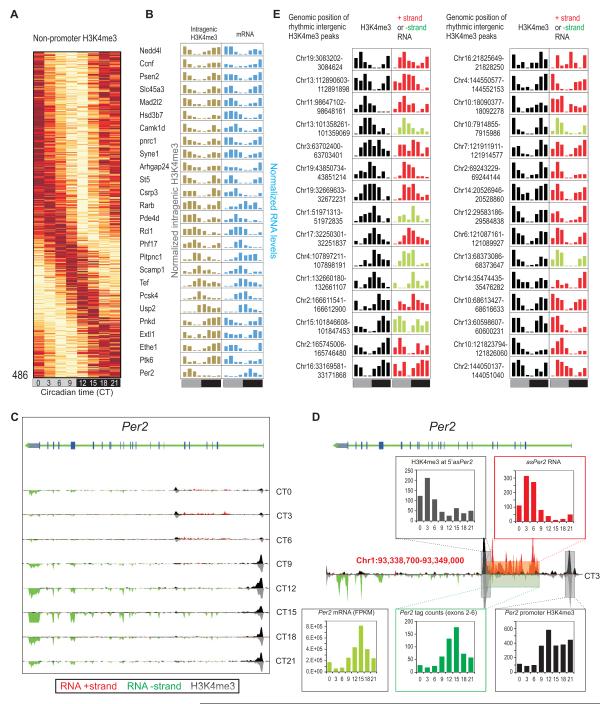
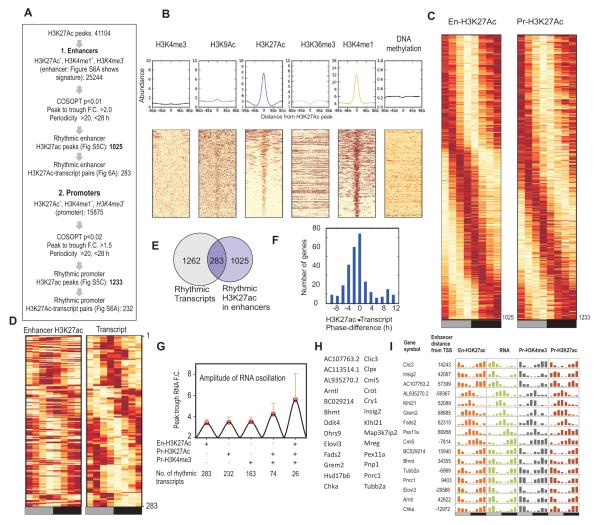
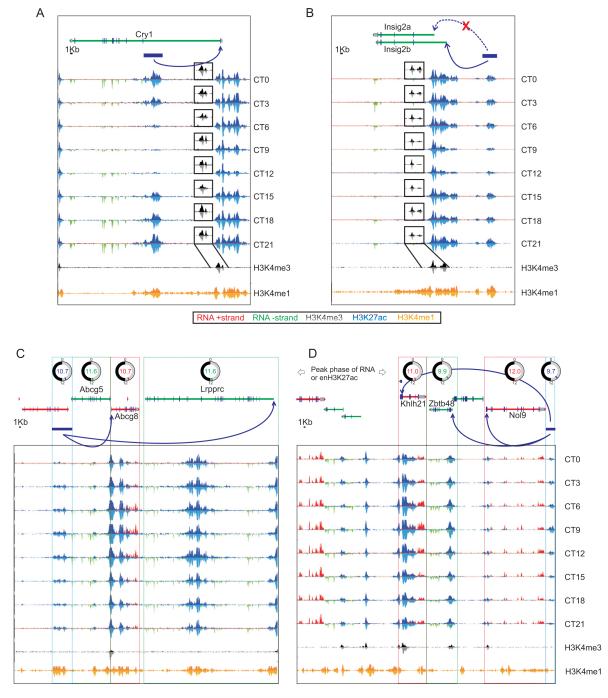
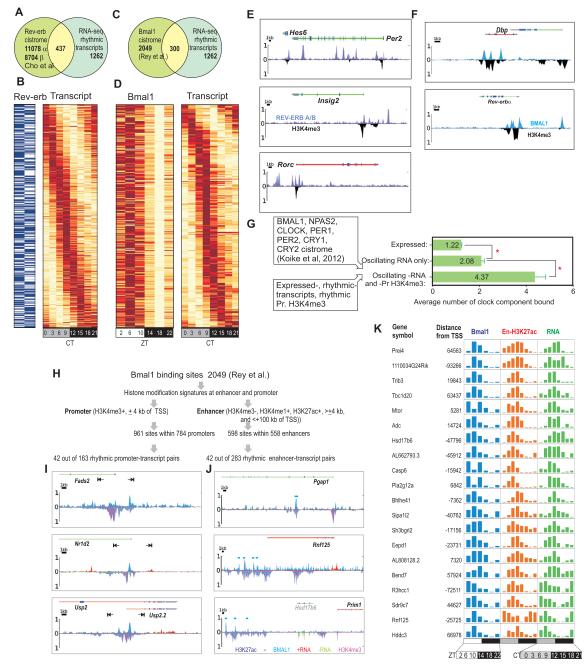
In the mouse liver, circadian transcriptional rhythms are necessary for metabolic homeostasis. Whether dynamic epigenomic modifications are associated with transcript oscillations has not been systematically investigated. We found that several antisense RNA, lincRNA, and microRNA transcripts also showed circadian oscillations in adult mouse livers. Robust transcript oscillations often correlated with rhythmic histone modifications in promoters, gene bodies, or enhancers, although promoter DNA methylation levels were relatively stable. Such integrative analyses identified oscillating expression of an antisense transcript (asPer2) to the gene encoding the circadian oscillator component Per2. Robust transcript oscillations often accompanied rhythms in multiple histone modifications and recruitment of multiple chromatin-associated clock components. Coupling of cycling histone modifications with nearby oscillating transcripts thus established a temporal relationship between enhancers, genes, and transcripts on a genome-wide scale in a mammalian liver. The results offer a framework for understanding the dynamics of metabolism, circadian clock, and chromatin modifications involved in metabolic homeostasis.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Anzulovich A, Mir A, Brewer M, Ferreyra G, Vinson C, Baler R. Elovl3: a model gene to dissect homeostatic links between the circadian clock and nutritional status. J Lipid Res. 2006;47:2690–2700. - PubMed
-
- Asher G, Schibler U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell metabolism. 2011;13:125–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
