

An interdomain energetic tug-of-war creates the allosterically active state in Hsp70 molecular chaperones
- PMID: 23217711
- PMCID: PMC3521165
- DOI: 10.1016/j.cell.2012.11.002
An interdomain energetic tug-of-war creates the allosterically active state in Hsp70 molecular chaperones
Abstract
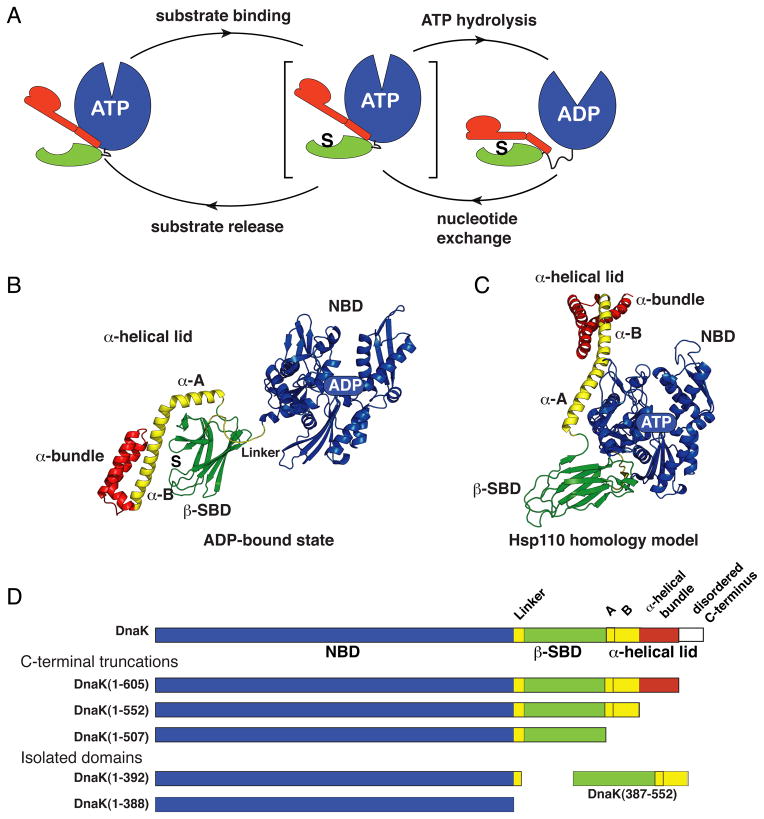
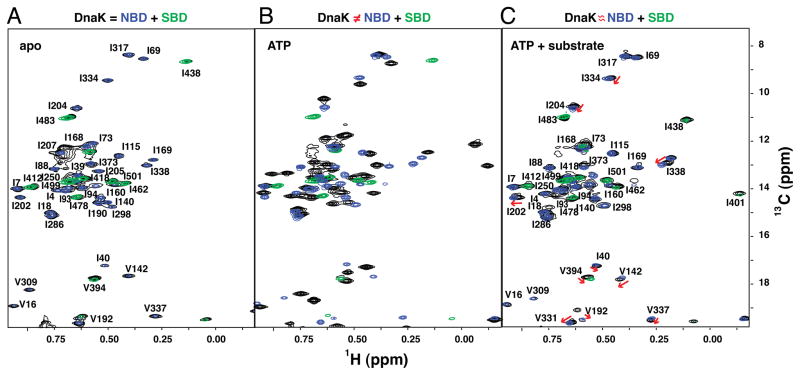
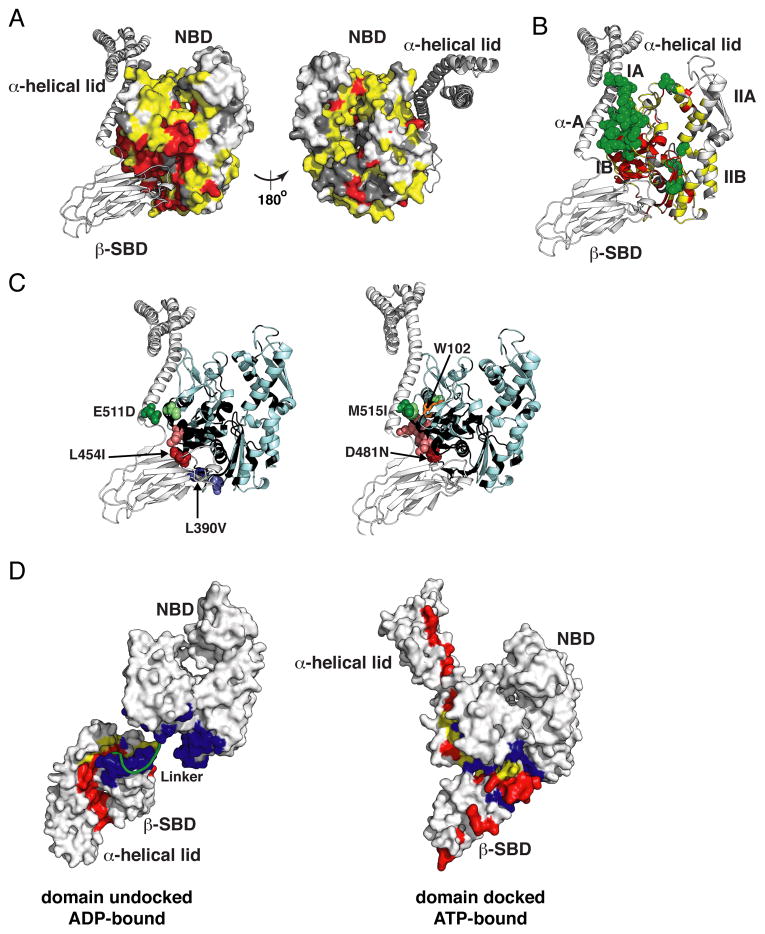
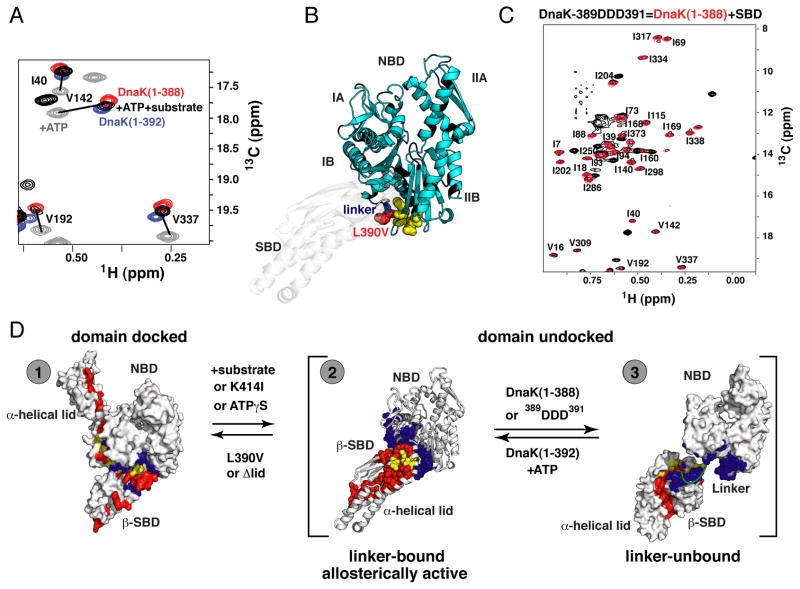
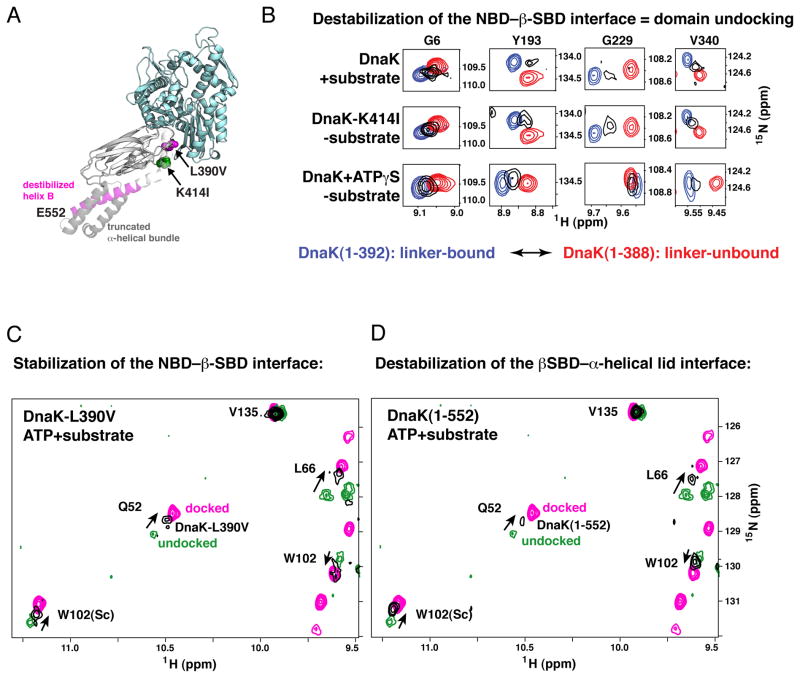
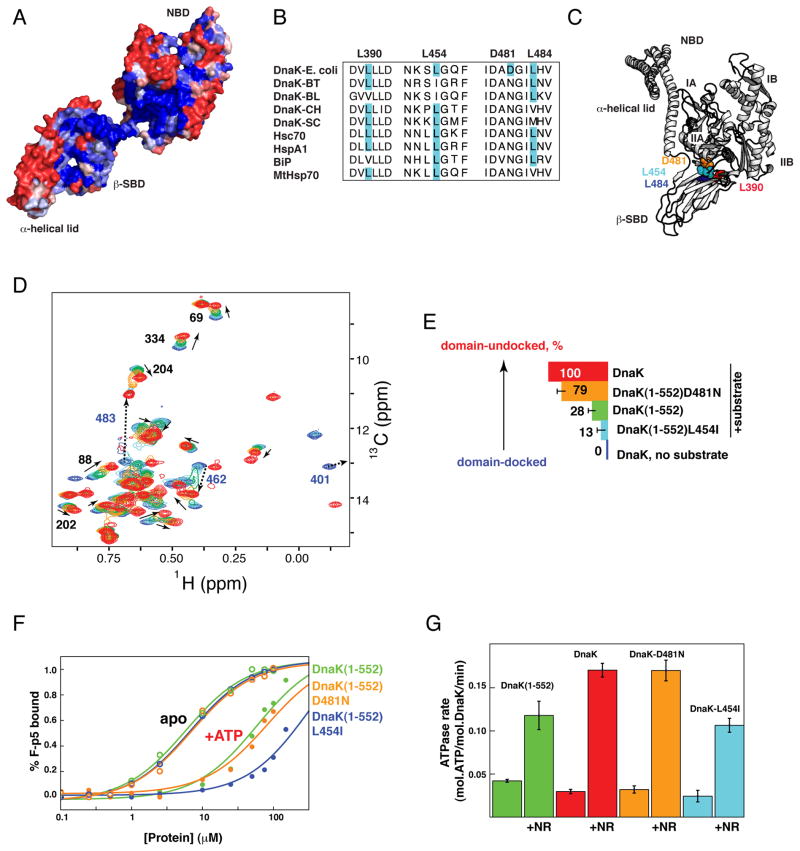
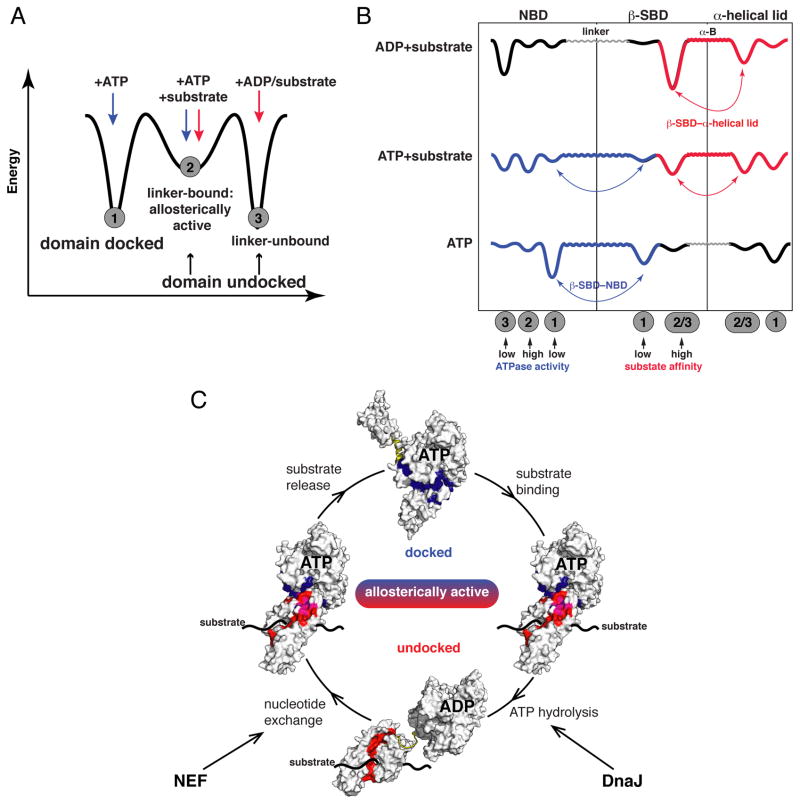
The allosteric mechanism of Hsp70 molecular chaperones enables ATP binding to the N-terminal nucleotide-binding domain (NBD) to alter substrate affinity to the C-terminal substrate-binding domain (SBD) and substrate binding to enhance ATP hydrolysis. Cycling between ATP-bound and ADP/substrate-bound states requires Hsp70s to visit a state with high ATPase activity and fast on/off kinetics of substrate binding. We have trapped this "allosterically active" state for the E. coli Hsp70, DnaK, and identified how interactions among the NBD, the β subdomain of the SBD, the SBD α-helical lid, and the conserved hydrophobic interdomain linker enable allosteric signal transmission between ligand-binding sites. Allostery in Hsp70s results from an energetic tug-of-war between domain conformations and formation of two orthogonal interfaces: between the NBD and SBD, and between the helical lid and the β subdomain of the SBD. The resulting energetic tension underlies Hsp70 functional properties and enables them to be modulated by ligands and cochaperones and "tuned" through evolution.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Allosteric opening of the polypeptide-binding site when an Hsp70 binds ATP.Nat Struct Mol Biol. 2013 Jul;20(7):900-7. doi: 10.1038/nsmb.2583. Epub 2013 May 26. Nat Struct Mol Biol. 2013. PMID: 23708608 Free PMC article.
-
Allosteric signal transmission in the nucleotide-binding domain of 70-kDa heat shock protein (Hsp70) molecular chaperones.Proc Natl Acad Sci U S A. 2011 Apr 26;108(17):6987-92. doi: 10.1073/pnas.1014448108. Epub 2011 Apr 11. Proc Natl Acad Sci U S A. 2011. PMID: 21482798 Free PMC article.
-
Substrate-binding domain conformational dynamics mediate Hsp70 allostery.Proc Natl Acad Sci U S A. 2015 Jun 2;112(22):E2865-73. doi: 10.1073/pnas.1506692112. Epub 2015 May 18. Proc Natl Acad Sci U S A. 2015. PMID: 26038563 Free PMC article.
-
The Link That Binds: The Linker of Hsp70 as a Helm of the Protein's Function.Biomolecules. 2019 Sep 27;9(10):543. doi: 10.3390/biom9100543. Biomolecules. 2019. PMID: 31569820 Free PMC article. Review.
-
Intra-molecular pathways of allosteric control in Hsp70s.Philos Trans R Soc Lond B Biol Sci. 2018 Jun 19;373(1749):20170183. doi: 10.1098/rstb.2017.0183. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 29735737 Free PMC article. Review.
Cited by
-
The Hsp70 chaperone system: distinct roles in erythrocyte formation and maintenance.Haematologica. 2021 Jun 1;106(6):1519-1534. doi: 10.3324/haematol.2019.233056. Haematologica. 2021. PMID: 33832207 Free PMC article. Review.
-
Hsp70 forms antiparallel dimers stabilized by post-translational modifications to position clients for transfer to Hsp90.Cell Rep. 2015 May 5;11(5):759-69. doi: 10.1016/j.celrep.2015.03.063. Epub 2015 Apr 23. Cell Rep. 2015. PMID: 25921532 Free PMC article.
-
Divide and conquer is always best: sensitivity of methyl correlation experiments.J Biomol NMR. 2013 Aug;56(4):331-5. doi: 10.1007/s10858-013-9751-9. Epub 2013 Jun 15. J Biomol NMR. 2013. PMID: 23771591 Free PMC article.
-
eIF1A/eIF5B interaction network and its functions in translation initiation complex assembly and remodeling.Nucleic Acids Res. 2016 Sep 6;44(15):7441-56. doi: 10.1093/nar/gkw552. Epub 2016 Jun 20. Nucleic Acids Res. 2016. PMID: 27325746 Free PMC article.
-
Hsp90 and Hsp70 chaperones: Collaborators in protein remodeling.J Biol Chem. 2019 Feb 8;294(6):2109-2120. doi: 10.1074/jbc.REV118.002806. Epub 2018 Nov 6. J Biol Chem. 2019. PMID: 30401745 Free PMC article. Review.
References
-
- Buchberger A, Theyssen H, Schroder H, McCarty JS, Virgallita G, Milkereit P, Reinstein J, Bukau B. Nucleotide-induced conformational-changes in the ATPase and substrate-binding domains of the DnaK chaperone provide evidence for interdomain communication. J Biol Chem. 1995;270:16903–16910. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
