

Crystallographic snapshot of cellulose synthesis and membrane translocation
- PMID: 23222542
- PMCID: PMC3542415
- DOI: 10.1038/nature11744
Crystallographic snapshot of cellulose synthesis and membrane translocation
Abstract
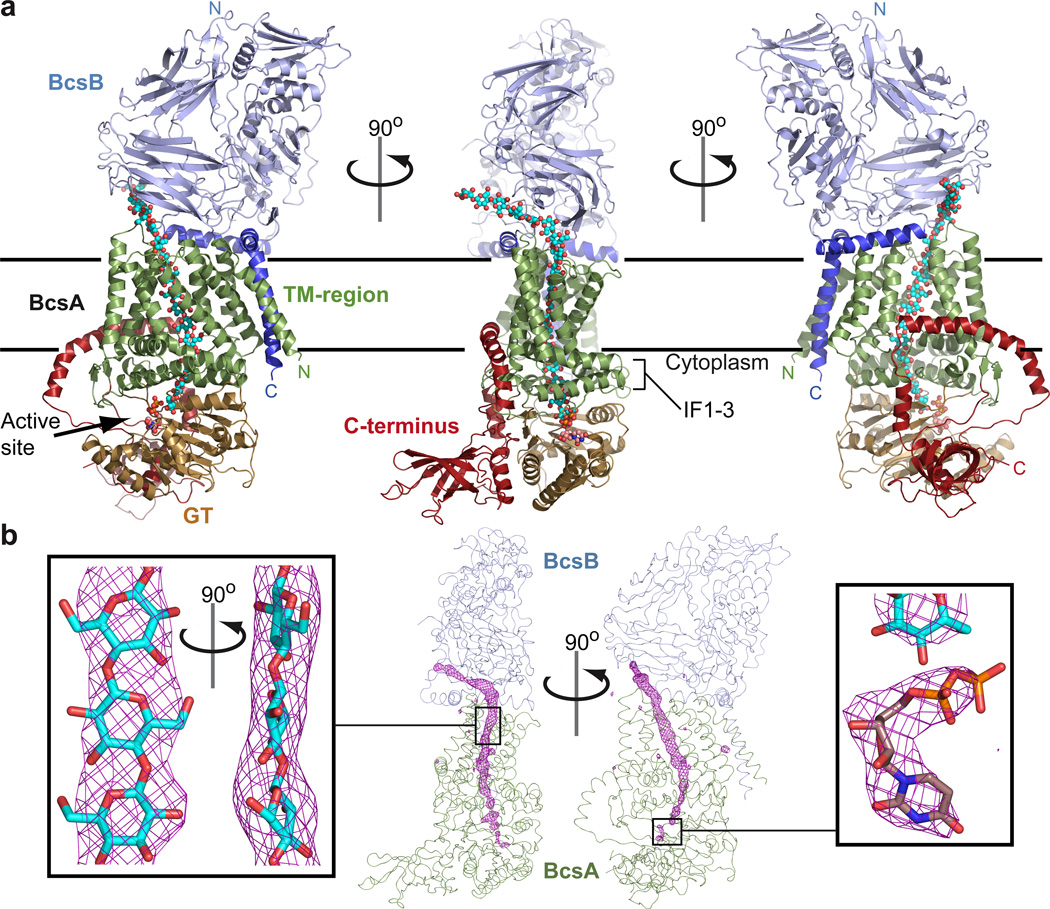
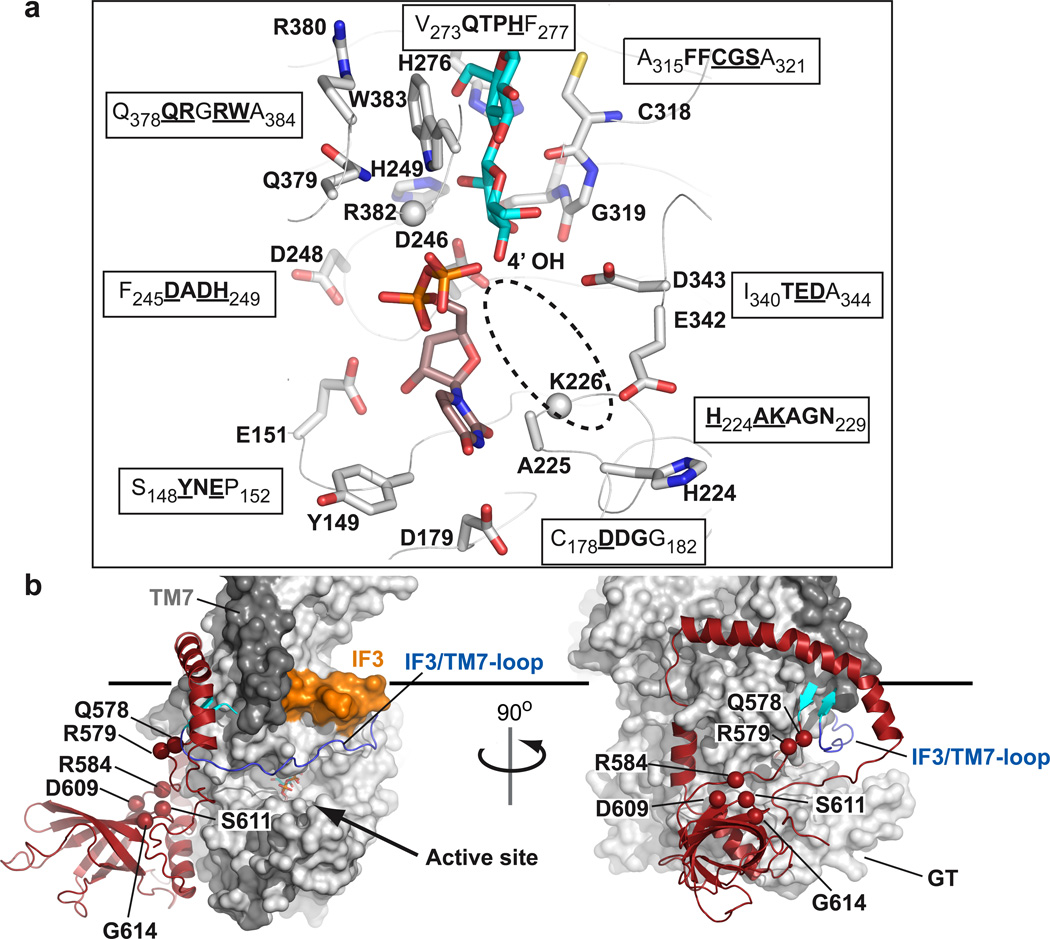
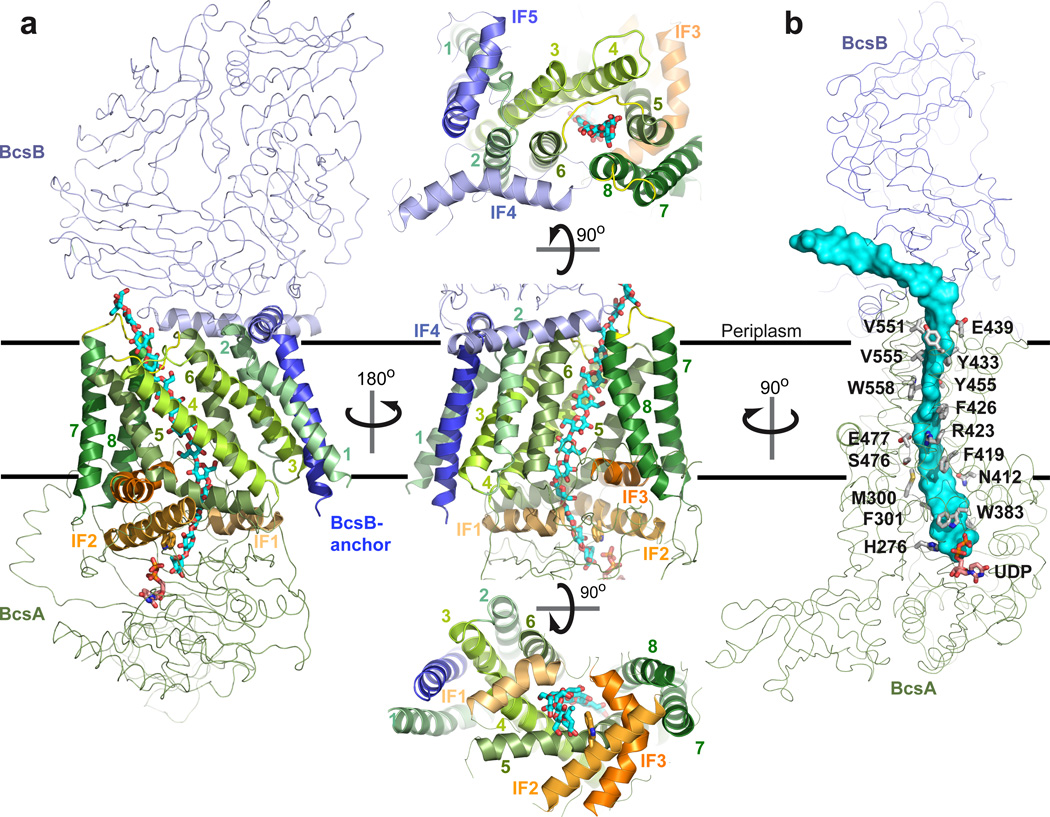
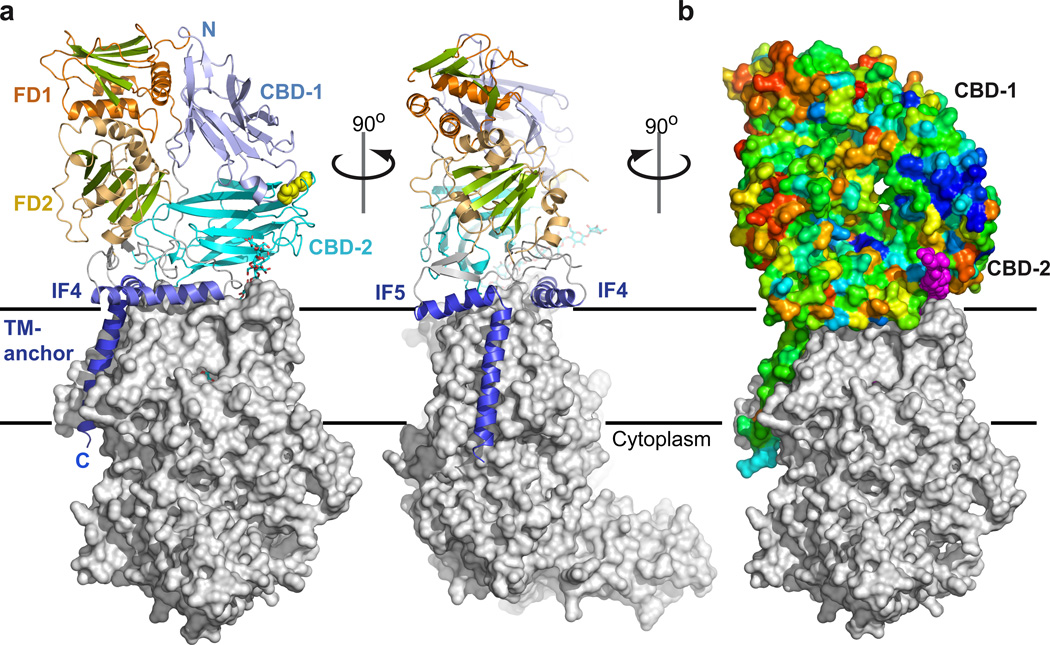
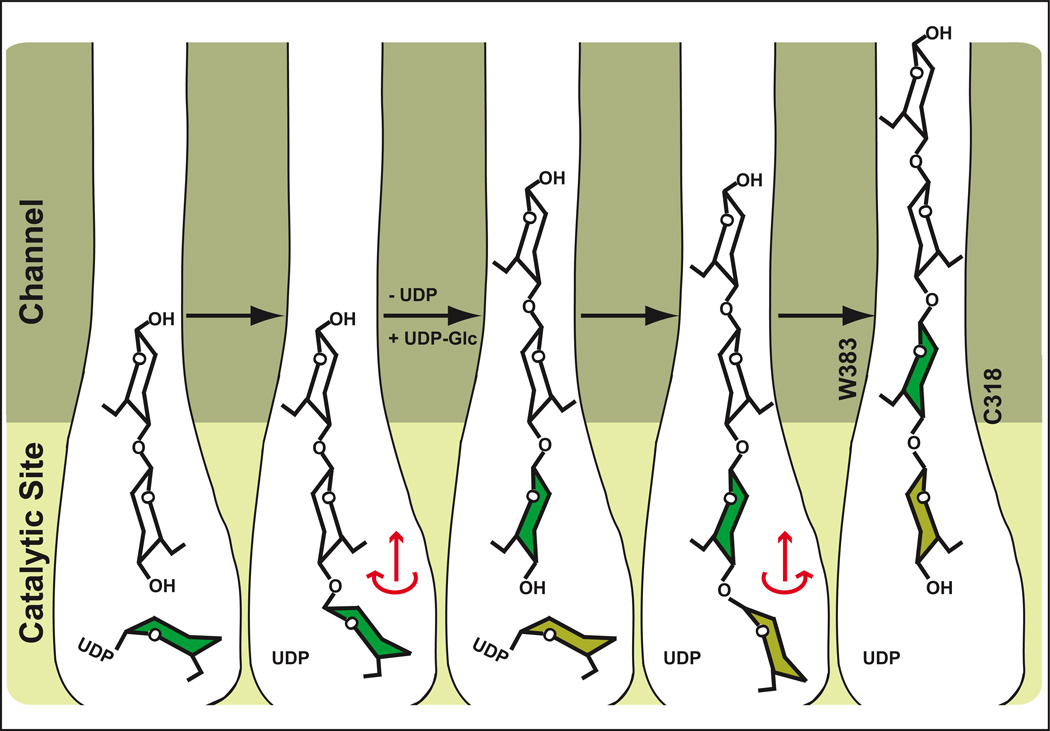
Cellulose, the most abundant biological macromolecule, is an extracellular, linear polymer of glucose molecules. It represents an essential component of plant cell walls but is also found in algae and bacteria. In bacteria, cellulose production frequently correlates with the formation of biofilms, a sessile, multicellular growth form. Cellulose synthesis and transport across the inner bacterial membrane is mediated by a complex of the membrane-integrated catalytic BcsA subunit and the membrane-anchored, periplasmic BcsB protein. Here we present the crystal structure of a complex of BcsA and BcsB from Rhodobacter sphaeroides containing a translocating polysaccharide. The structure of the BcsA-BcsB translocation intermediate reveals the architecture of the cellulose synthase, demonstrates how BcsA forms a cellulose-conducting channel, and suggests a model for the coupling of cellulose synthesis and translocation in which the nascent polysaccharide is extended by one glucose molecule at a time.
Figures
References
-
- Somerville C. Cellulose synthesis in higher plants. Annu Rev Cell Dev Biol. 2006;22:53–78. - PubMed
-
- Merzendorfer H. Insect chitin synthases: a review. J Comp Physiol B, Biochem Syst Environ Physiol. 2006;176:1–15. - PubMed
-
- Hubbard C, McNamara JT, Azumaya C, Patel MS, Zimmer J. The hyaluronan synthase catalyzes the synthesis and membrane translocation of hyaluronan. J Mol Biol. 2012;418:21–31. - PubMed
-
- Nishiyama Y, Langan P, Chanzy H. Crystal structure and hydrogen-bonding system in cellulose Ibeta from synchrotron X-ray and neutron fiber diffraction. J Am Chem Soc. 2002;124:9074–9082. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
