

Nondisjunctional segregations in Drosophila female meiosis I are preceded by homolog malorientation at metaphase arrest
- PMID: 23222652
- PMCID: PMC3567735
- DOI: 10.1534/genetics.112.146241
Nondisjunctional segregations in Drosophila female meiosis I are preceded by homolog malorientation at metaphase arrest
Abstract
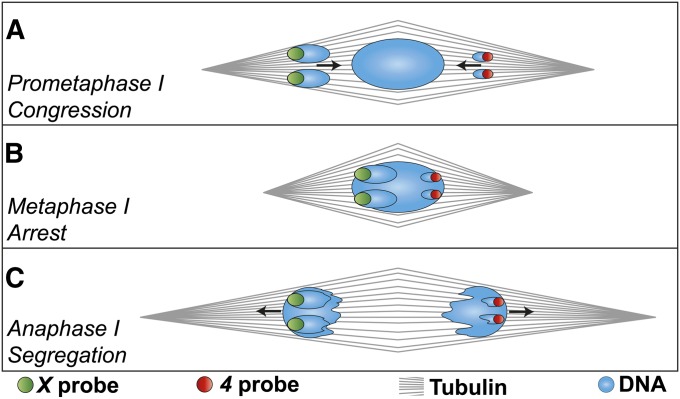
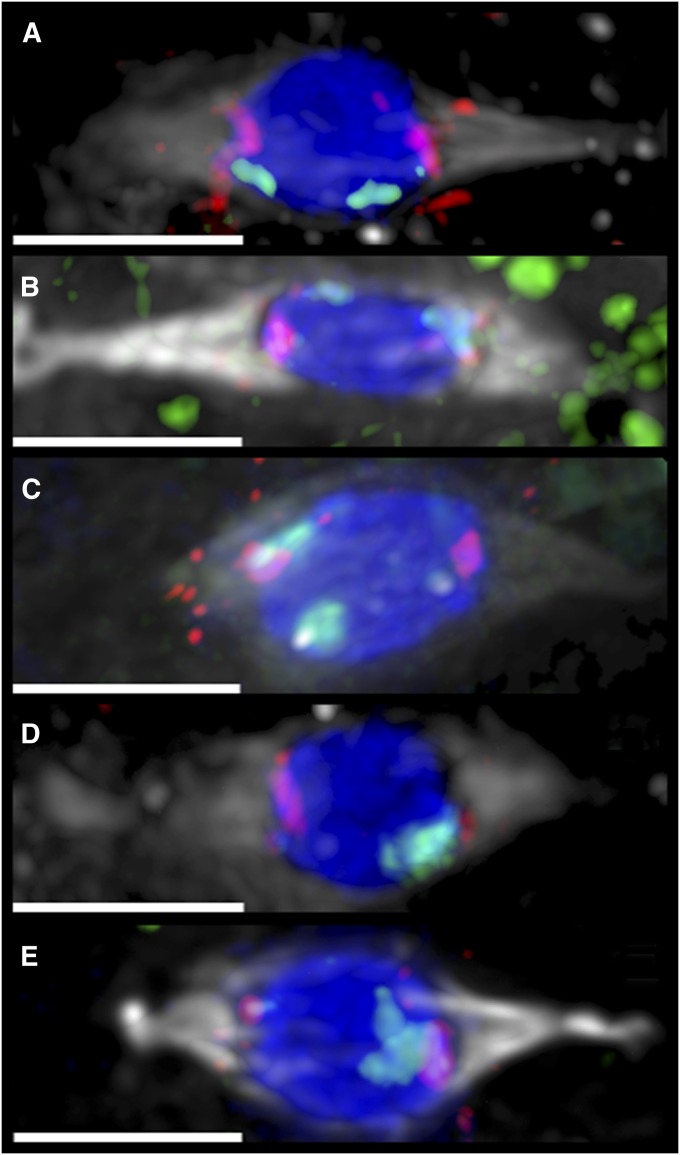
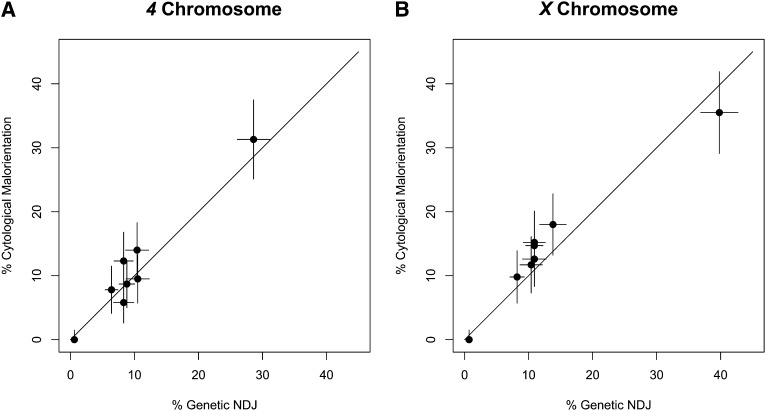
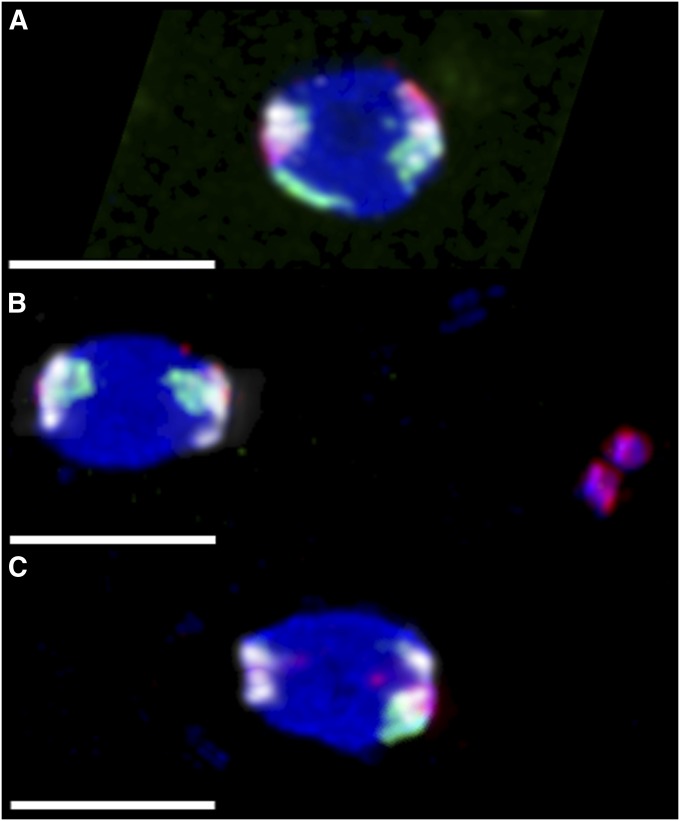
The model of Drosophila female meiosis I was recently revised by the discovery that chromosome congression precedes metaphase I arrest. Use of the prior framework to interpret data from meiotic mutants led to the conclusion that chromosome segregation errors (nondisjunction, NDJ) occurred when nonexchange chromosomes moved out on the spindle in a maloriented configuration and became trapped there at metaphase arrest. The discovery that congression returns nonexchange chromosomes to the metaphase plate invalidates this interpretation and raises the question of what events actually do lead to NDJ. To address this, we have assayed an allelic series of ald (mps1) meiotic mutants that complete congression at wild-type rates, but have widely varying NDJ rates in an otherwise isogenic background, as well as a nod mutant background that primarily undergoes loss of chromosome 4. Using genetic assays to measure NDJ rates, and FISH assays to measure chromosome malorientation rates in metaphase-arrested oocytes, shows that these two rates are highly correlated across ald mutants, suggesting that malorientation during congression commits these chromosomes to eventually nondisjoin. Likewise, the rate of chromosome loss observed in nod is similar to the rate at which these chromosomes fail to associate with the main chromosome mass. Together these results provide a proximal mechanism for how these meiotic mutants cause NDJ and chromosome loss and improve our understanding of how prometaphase chromosome congression relates to anaphase chromosome segregation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
