

Proline-dependent regulation of Clostridium difficile Stickland metabolism
- PMID: 23222730
- PMCID: PMC3562115
- DOI: 10.1128/JB.01492-12
Proline-dependent regulation of Clostridium difficile Stickland metabolism
Abstract
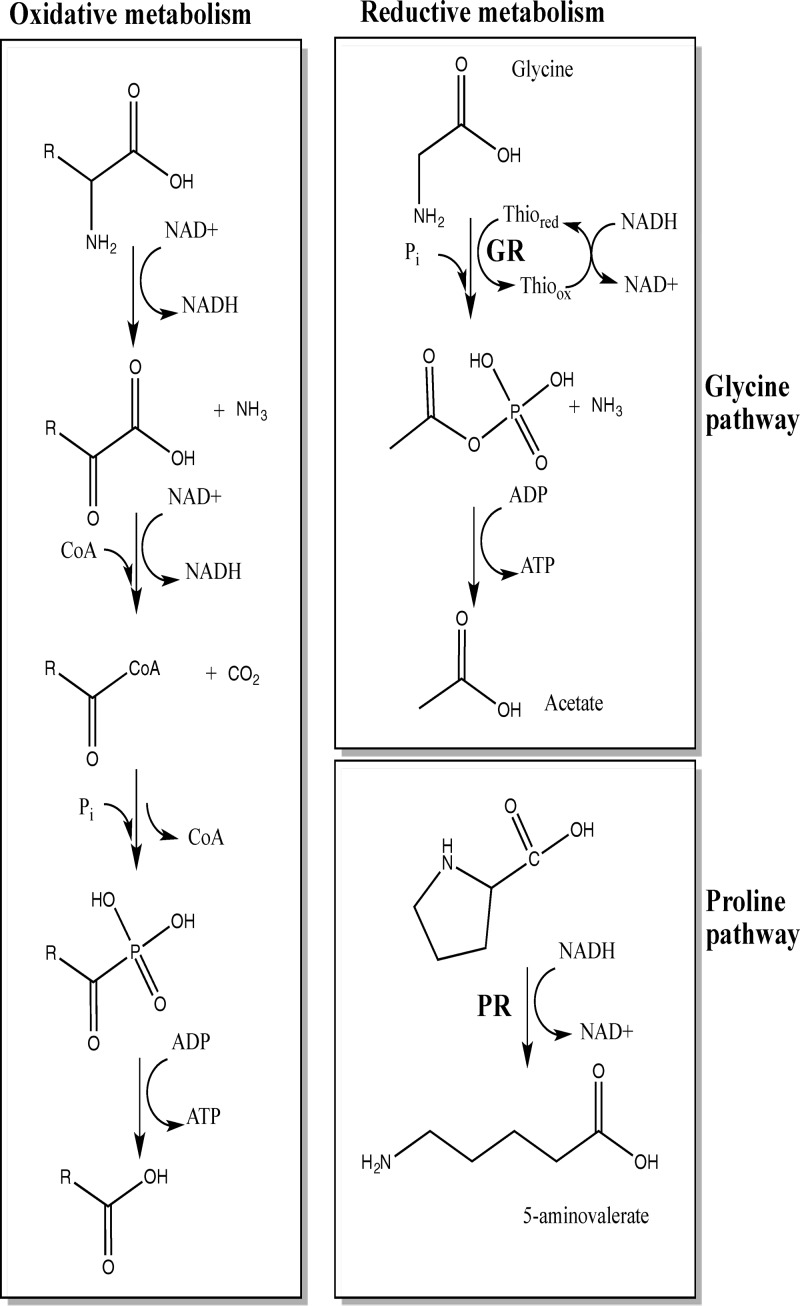
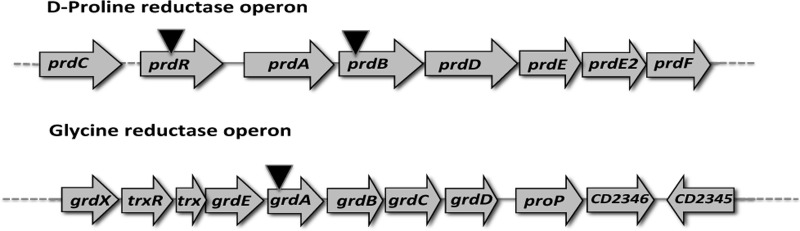
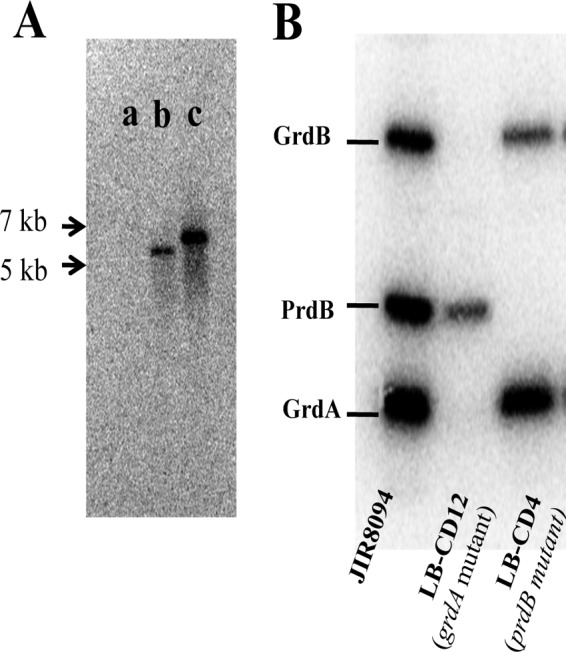
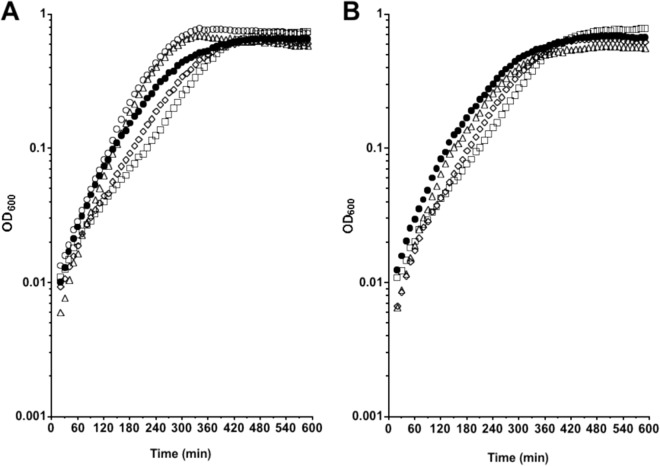
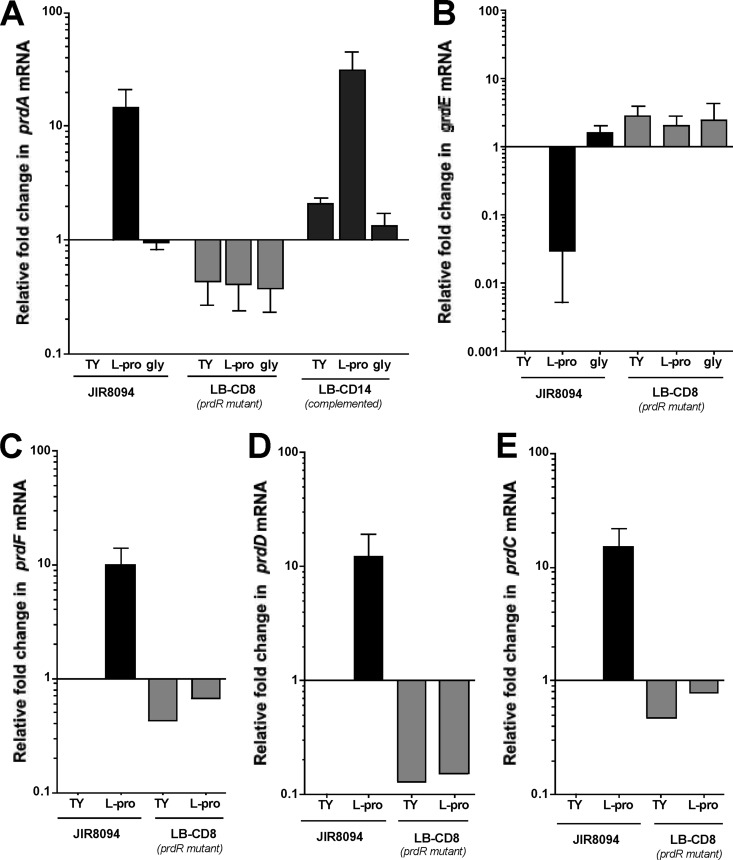
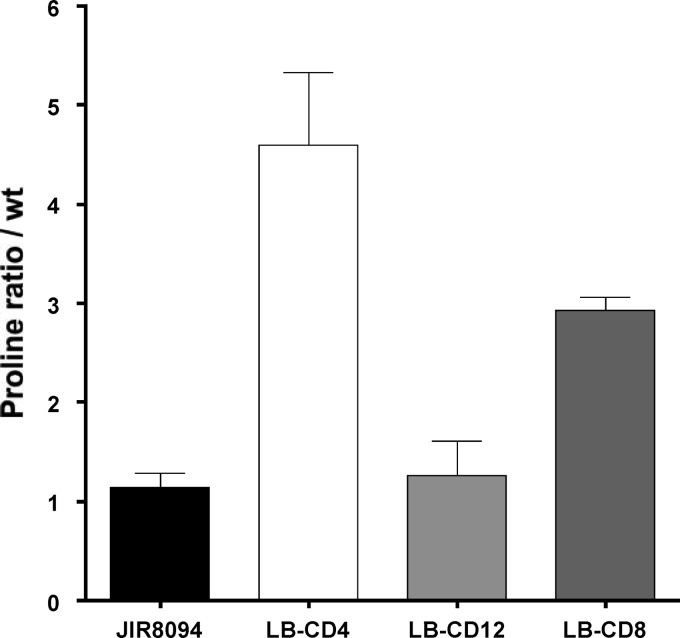
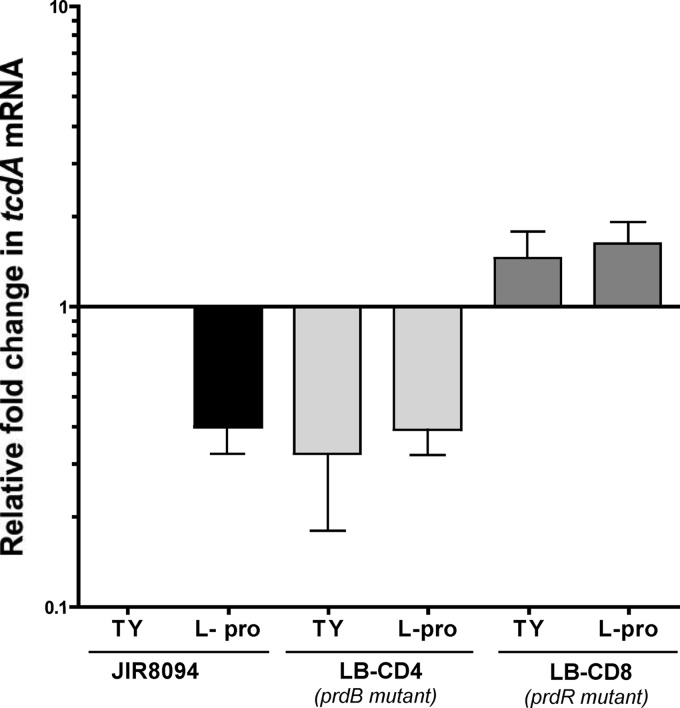
Clostridium difficile, a proteolytic Gram-positive anaerobe, has emerged as a significant nosocomial pathogen. Stickland fermentation reactions are thought to be important for growth of C. difficile and appear to influence toxin production. In Stickland reactions, pairs of amino acids donate and accept electrons, generating ATP and reducing power in the process. Reduction of the electron acceptors proline and glycine requires the d-proline reductase (PR) and the glycine reductase (GR) enzyme complexes, respectively. Addition of proline in the medium increases the level of PR protein but decreases the level of GR. We report the identification of PrdR, a protein that activates transcription of the PR-encoding genes in the presence of proline and negatively regulates the GR-encoding genes. The results suggest that PrdR is a central metabolism regulator that controls preferential utilization of proline and glycine to produce energy via the Stickland reactions.
Figures

References
-
- Johnson S, Clabots CR, Linn FV, Olson MM, Peterson LR, Gerding DN. 1990. Nosocomial Clostridium difficile colonisation and disease. Lancet 336:97–100 - PubMed
-
- Kelly CP, LaMont JT. 2008. Clostridium difficile—more difficult than ever. N. Engl. J. Med. 359:1932–1940 - PubMed
-
- Warny M, Pepin J, Fang A, Killgore G, Thompson A, Brazier J, Frost E, McDonald LC. 2005. Toxin production by an emerging strain of Clostridium difficile associated with outbreaks of severe disease in North America and Europe. Lancet 366:1079–1084 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
