

Deletion of tetraspanin CD9 diminishes lymphangiogenesis in vivo and in vitro
- PMID: 23223239
- PMCID: PMC3554885
- DOI: 10.1074/jbc.M112.424291
Deletion of tetraspanin CD9 diminishes lymphangiogenesis in vivo and in vitro
Abstract
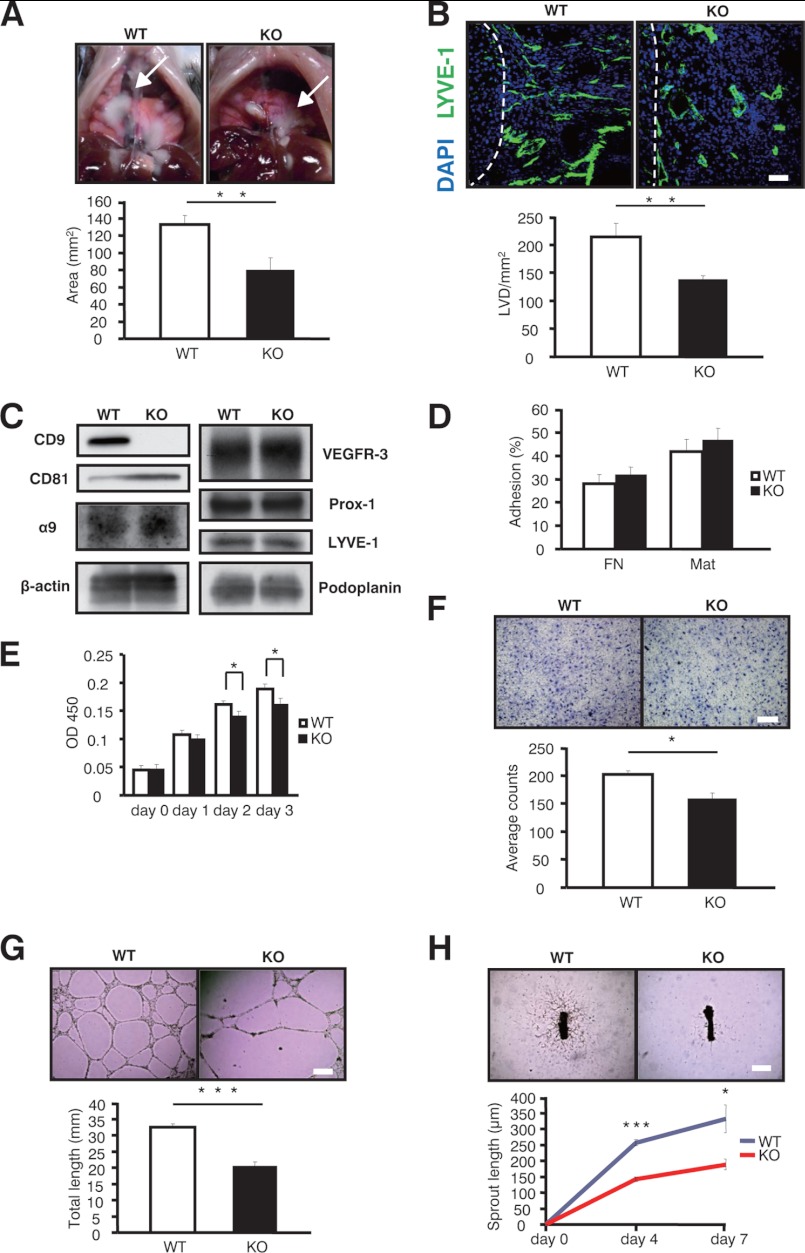
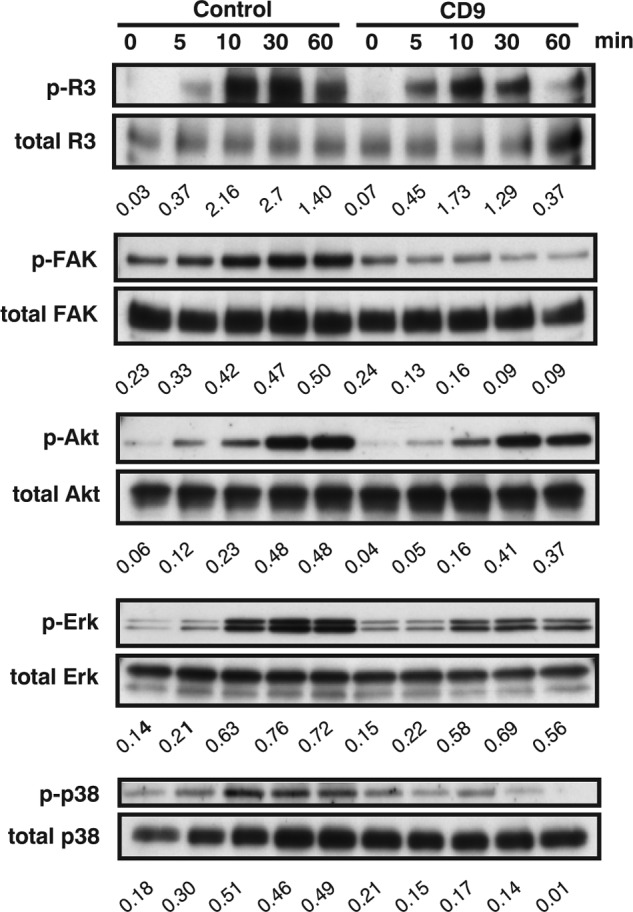
Tetraspanins have emerged as key players in malignancy and inflammatory diseases, yet little is known about their roles in angiogenesis, and nothing is known about their involvement in lymphangiogenesis. We found here that tetraspanins are abundantly expressed in human lymphatic endothelial cells (LEC). After intrathoracic tumor implantation, metastasis to lymph nodes was diminished and accompanied by decreased angiogenesis and lymphangiogenesis in tetraspanin CD9-KO mice. Moreover, lymphangiomas induced in CD9-KO mice were less pronounced with decreased lymphangiogenesis compared with those in wild-type mice. Although mouse LEC isolated from CD9-KO mice showed normal adhesion, lymphangiogenesis was markedly impaired in several assays (migration, proliferation, and cable formation) in vitro and in the lymphatic ring assay ex vivo. Consistent with these findings in mouse LEC, knocking down CD9 in human LEC also produced decreased migration, proliferation, and cable formation. Immunoprecipitation analysis demonstrated that deletion of CD9 in LEC diminished formation of functional complexes between VEGF receptor-3 and integrins (α5 and α9). Therefore, knocking down CD9 in LEC attenuated VEGF receptor-3 signaling, as well as downstream signaling such as Erk and p38 upon VEGF-C stimulation. Finally, double deletion of CD9/CD81 in mice caused abnormal development of lymphatic vasculature in the trachea and diaphragm, suggesting that CD9 and a closely related tetraspanin CD81 coordinately play an essential role in physiological lymphangiogenesis. In conclusion, tetraspanin CD9 modulates molecular organization of integrins in LEC, thereby supporting several functions required for lymphangiogenesis.
Figures








References
-
- Adams R. H., Alitalo K. (2007) Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 8, 464–478 - PubMed
-
- Grothey A., Galanis E. (2009) Targeting angiogenesis: progress with anti-VEGF treatment with large molecules. Nat. Rev. Clin. Oncol. 6, 507–518 - PubMed
-
- Tammela T., Alitalo K. (2010) Lymphangiogenesis. Molecular mechanisms and future promise. Cell 140, 460–476 - PubMed
-
- Alitalo K., Tammela T., Petrova T. V. (2005) Lymphangiogenesis in development and human disease. Nature 438, 946–953 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
