

Unique thrombin inhibition mechanism by anophelin, an anticoagulant from the malaria vector
- PMID: 23223529
- PMCID: PMC3535606
- DOI: 10.1073/pnas.1211614109
Unique thrombin inhibition mechanism by anophelin, an anticoagulant from the malaria vector
Abstract
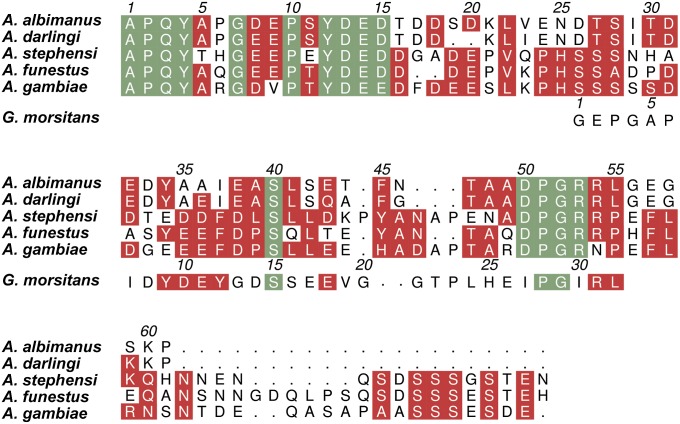
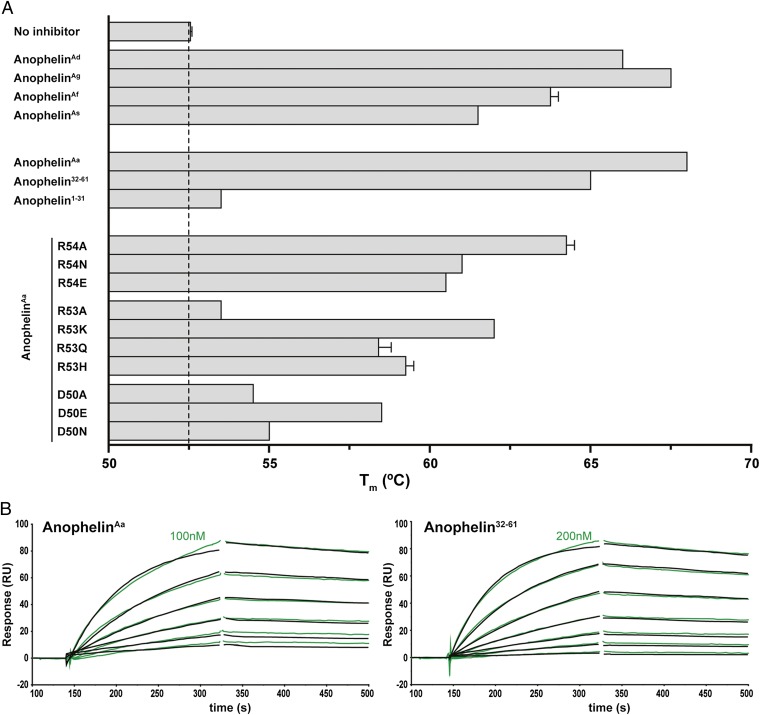
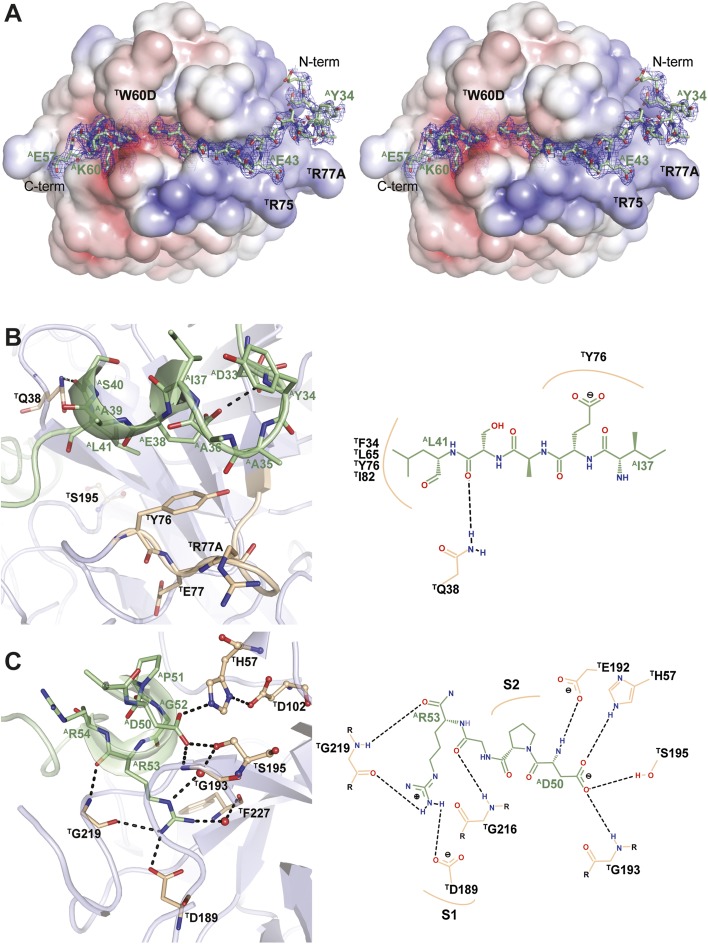
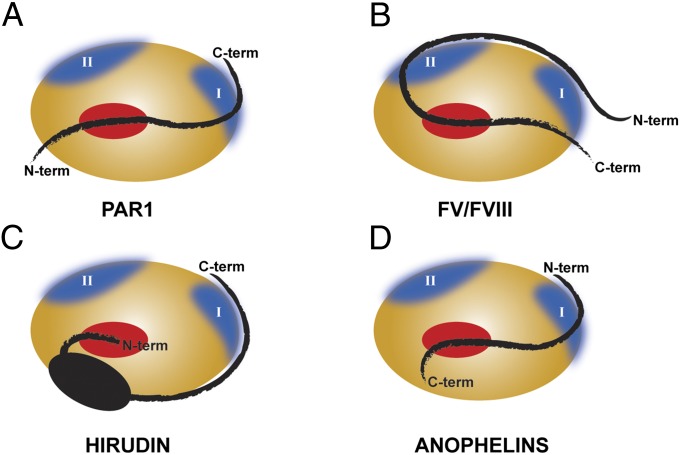
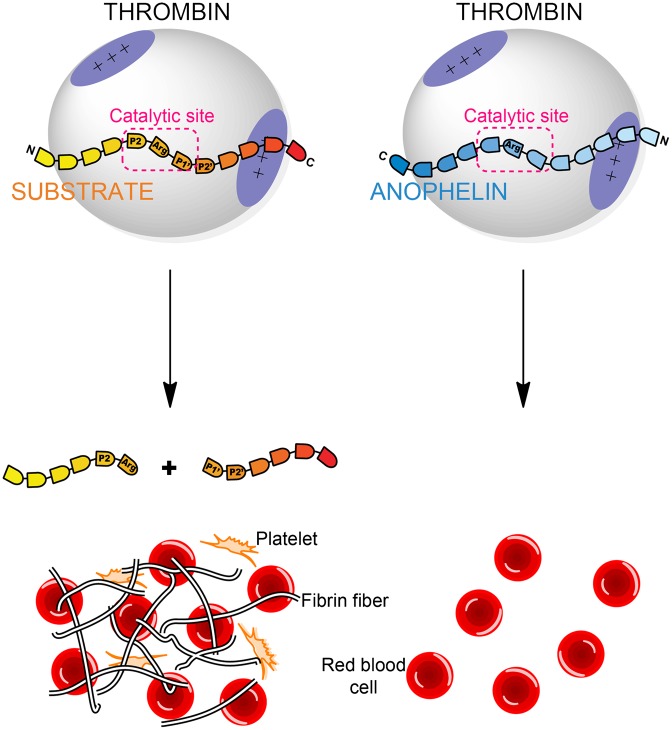
Anopheles mosquitoes are vectors of malaria, a potentially fatal blood disease affecting half a billion humans worldwide. These blood-feeding insects include in their antihemostatic arsenal a potent thrombin inhibitor, the flexible and cysteine-less anophelin. Here, we present a thorough structure-and-function analysis of thrombin inhibition by anophelin, including the 2.3-Å crystal structure of the human thrombin·anophelin complex. Anophelin residues 32-61 are well-defined by electron density, completely occupying the long cleft between the active site and exosite I. However, in striking contrast to substrates, the D50-R53 anophelin tetrapeptide occupies the active site cleft of the enzyme, whereas the upstream residues A35-P45 shield the regulatory exosite I, defining a unique reverse-binding mode of an inhibitor to the target proteinase. The extensive interactions established, the disruption of thrombin's active site charge-relay system, and the insertion of residue R53 into the proteinase S(1) pocket in an orientation opposed to productive substrates explain anophelin's remarkable specificity and resistance to proteolysis by thrombin. Complementary biophysical and functional characterization of point mutants and truncated versions of anophelin unambiguously establish the molecular mechanism of action of this family of serine proteinase inhibitors (I77). These findings have implications for the design of novel antithrombotics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Functional analyses yield detailed insight into the mechanism of thrombin inhibition by the antihemostatic salivary protein cE5 from Anopheles gambiae.J Biol Chem. 2017 Jul 28;292(30):12632-12642. doi: 10.1074/jbc.M117.788042. Epub 2017 Jun 7. J Biol Chem. 2017. PMID: 28592490 Free PMC article.
-
Anophelin: kinetics and mechanism of thrombin inhibition.Biochemistry. 1999 Dec 14;38(50):16678-85. doi: 10.1021/bi991231p. Biochemistry. 1999. PMID: 10600131
-
Identification and characterization of novel salivary thrombin inhibitors from the ixodidae tick, Haemaphysalis longicornis.Eur J Biochem. 2003 May;270(9):1926-34. doi: 10.1046/j.1432-1033.2003.03560.x. Eur J Biochem. 2003. PMID: 12709051
-
Thrombin inhibitors as antithrombotic agents: the importance of rapid inhibition.J Enzyme Inhib. 1995;9(1):3-15. doi: 10.3109/14756369509040677. J Enzyme Inhib. 1995. PMID: 8568565 Review.
-
The Research Progress of Direct Thrombin Inhibitors.Mini Rev Med Chem. 2020;20(16):1574-1585. doi: 10.2174/1389557519666191015201125. Mini Rev Med Chem. 2020. PMID: 31644402 Review.
Cited by
-
Rapid assembly and profiling of an anticoagulant sulfoprotein library.Proc Natl Acad Sci U S A. 2019 Jul 9;116(28):13873-13878. doi: 10.1073/pnas.1905177116. Epub 2019 Jun 20. Proc Natl Acad Sci U S A. 2019. PMID: 31221752 Free PMC article.
-
Unraveling dual feeding associated molecular complexity of salivary glands in the mosquito Anopheles culicifacies.Biol Open. 2015 Jul 10;4(8):1002-15. doi: 10.1242/bio.012294. Biol Open. 2015. PMID: 26163527 Free PMC article.
-
Anopheline salivary protein genes and gene families: an evolutionary overview after the whole genome sequence of sixteen Anopheles species.BMC Genomics. 2017 Feb 13;18(1):153. doi: 10.1186/s12864-017-3579-8. BMC Genomics. 2017. PMID: 28193177 Free PMC article.
-
The tick-derived anticoagulant madanin is processed by thrombin and factor Xa.PLoS One. 2013 Aug 12;8(8):e71866. doi: 10.1371/journal.pone.0071866. eCollection 2013. PLoS One. 2013. PMID: 23951260 Free PMC article.
-
Salivary Digestion Extends the Range of Sugar-Aversions in the German Cockroach.Insects. 2021 Mar 21;12(3):263. doi: 10.3390/insects12030263. Insects. 2021. PMID: 33801079 Free PMC article.
References
-
- Valenzuela JG, Francischetti IMB, Pham VM, Garfield MK, Ribeiro JMC. Exploring the salivary gland transcriptome and proteome of the Anopheles stephensi mosquito. Insect Biochem Mol Biol. 2003;33(7):717–732. - PubMed
-
- World Health Organization . World Malaria Report. Geneva: World Health Organization; 2010.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
