

A novel Rac1 GAP splice variant relays poly-Ub accumulation signals to mediate Rac1 inactivation
- PMID: 23223568
- PMCID: PMC3564530
- DOI: 10.1091/mbc.E12-07-0565
A novel Rac1 GAP splice variant relays poly-Ub accumulation signals to mediate Rac1 inactivation
Abstract
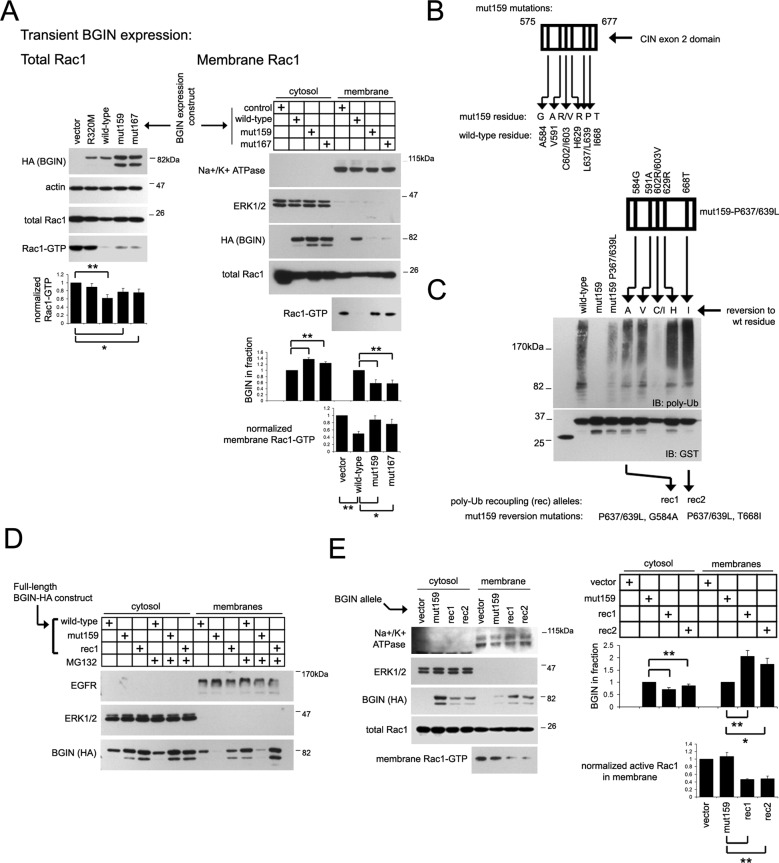
Spatial control of RhoGTPase-inactivating GAP components remains largely enigmatic. We describe a brain-specific RhoGAP splice variant, BARGIN (BGIN), which comprises a combination of BAR, GAP, and partial CIN phosphatase domains spliced from adjacent SH3BP1 and CIN gene loci. Excision of BGIN exon 2 results in recoding of a 42-amino acid N-terminal stretch. The partial CIN domain is a poly-ubiquitin (poly-Ub)-binding module that facilitates BGIN distribution to membranous and detergent-insoluble fractions. Poly-Ub/BGIN interactions support BGIN-mediated inactivation of a membranous Rac1 population, which consequently inactivates membrane-localized Rac1 effector systems such as reactive oxygen species (ROS) generation by the Nox1 complex. Given that Ub aggregate pathology and proteotoxicity are central themes in various neurodegenerative disorders, we investigated whether BGIN/Rac1 signaling could be involved in neurodegenerative proteotoxicity. BGIN/Ub interactions are observed through colocalization in tangle aggregates in the Alzheimer's disease (AD) brain. Moreover, enhanced BGIN membrane distribution correlates with reduced Rac1 activity in AD brain tissue. Finally, BGIN contributes to Rac1 inhibition and ROS generation in an amyloid precursor protein (APP) proteotoxicity model. These results suggest that BGIN/poly-Ub interactions enhance BGIN membrane distribution and relay poly-Ub signals to enact Rac1 inactivation, and attenuation of Rac1 signaling is partially dependent on BGIN in a proteotoxic APP context.
Figures







Similar articles
-
Nox1-dependent reactive oxygen generation is regulated by Rac1.J Biol Chem. 2006 Jun 30;281(26):17718-26. doi: 10.1074/jbc.M512751200. Epub 2006 Apr 24. J Biol Chem. 2006. PMID: 16636067
-
Hace1 controls ROS generation of vertebrate Rac1-dependent NADPH oxidase complexes.Nat Commun. 2013;4:2180. doi: 10.1038/ncomms3180. Nat Commun. 2013. PMID: 23864022 Free PMC article.
-
A stretch of polybasic residues mediates Cdc42 GTPase-activating protein (CdGAP) binding to phosphatidylinositol 3,4,5-trisphosphate and regulates its GAP activity.J Biol Chem. 2012 Jun 1;287(23):19610-21. doi: 10.1074/jbc.M112.344606. Epub 2012 Apr 19. J Biol Chem. 2012. PMID: 22518840 Free PMC article.
-
Impaired autophagy and APP processing in Alzheimer's disease: The potential role of Beclin 1 interactome.Prog Neurobiol. 2013 Jul-Aug;106-107:33-54. doi: 10.1016/j.pneurobio.2013.06.002. Epub 2013 Jul 1. Prog Neurobiol. 2013. PMID: 23827971 Review.
-
Crosstalk between Rac1-mediated actin regulation and ROS production.Free Radic Biol Med. 2018 Feb 20;116:101-113. doi: 10.1016/j.freeradbiomed.2018.01.008. Epub 2018 Jan 10. Free Radic Biol Med. 2018. PMID: 29330095 Review.
Cited by
-
Inhibition of Rac1-dependent forgetting alleviates memory deficits in animal models of Alzheimer's disease.Protein Cell. 2019 Oct;10(10):745-759. doi: 10.1007/s13238-019-0641-0. Epub 2019 Jul 18. Protein Cell. 2019. PMID: 31321704 Free PMC article.
-
SH3-domain binding protein 1 in the tumor microenvironment promotes hepatocellular carcinoma metastasis through WAVE2 pathway.Oncotarget. 2016 Apr 5;7(14):18356-70. doi: 10.18632/oncotarget.7786. Oncotarget. 2016. PMID: 26933917 Free PMC article.
-
SORLA attenuates EphA4 signaling and amyloid β-induced neurodegeneration.J Exp Med. 2017 Dec 4;214(12):3669-3685. doi: 10.1084/jem.20171413. Epub 2017 Nov 7. J Exp Med. 2017. PMID: 29114064 Free PMC article.
-
Suppression of Rac1 Signaling by Influenza A Virus NS1 Facilitates Viral Replication.Sci Rep. 2016 Nov 21;6:35041. doi: 10.1038/srep35041. Sci Rep. 2016. PMID: 27869202 Free PMC article.
-
SNX27 and SORLA Interact to Reduce Amyloidogenic Subcellular Distribution and Processing of Amyloid Precursor Protein.J Neurosci. 2016 Jul 27;36(30):7996-8011. doi: 10.1523/JNEUROSCI.0206-16.2016. J Neurosci. 2016. PMID: 27466343 Free PMC article.
References
-
- Alves-Rodrigues A, Gregori L, Figueiredo-Pereira ME. Ubiquitin, cellular inclusions and their role in neurodegeneration. Trends Neurosci. 1998;21:516–520. - PubMed
-
- Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev. 2007;87:245–313. - PubMed
-
- Benard V, Bohl BP, Bokoch GM. Characterization of rac and cdc42 activation in chemoattractant-stimulated human neutrophils using a novel assay for active GTPases. J Biol Chem. 1999;274:13198–13204. - PubMed
-
- Billuart P, et al. Oligophrenin-1 encodes a rhoGAP protein involved in X-linked mental retardation. Nature. 1998;392:923–926. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P41 GM103533/GM/NIGMS NIH HHS/United States
- P50 AG005131/AG/NIA NIH HHS/United States
- 5P41RR011823-17/RR/NCRR NIH HHS/United States
- P30 NS076411/NS/NINDS NIH HHS/United States
- P01 AG031097/AG/NIA NIH HHS/United States
- R01AG027463-04/AG/NIA NIH HHS/United States
- R01 GM044428/GM/NIGMS NIH HHS/United States
- AG5131/AG/NIA NIH HHS/United States
- P41 RR011823/RR/NCRR NIH HHS/United States
- GM044428/GM/NIGMS NIH HHS/United States
- R01 AG018440/AG/NIA NIH HHS/United States
- R01 AG027463/AG/NIA NIH HHS/United States
- R37 AG018440/AG/NIA NIH HHS/United States
- P01 AG010435/AG/NIA NIH HHS/United States
- 8 P41 GM103533-17/GM/NIGMS NIH HHS/United States
- AG18440/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
