

Role of TGF-β signaling in curcumin-mediated inhibition of tumorigenicity of human lung cancer cells
- PMID: 23224523
- PMCID: PMC4295202
- DOI: 10.1007/s00432-012-1352-6
Role of TGF-β signaling in curcumin-mediated inhibition of tumorigenicity of human lung cancer cells
Abstract
Purpose: Curcumin has been shown to have potent anticancer activities like inhibition of cell proliferation, induction of apoptosis, and suppression of angiogenesis. Transforming growth factor-β (TGF-β) signaling plays a complex role in tumor suppression and promotion depending on the tumor type and stage. However, the effect of curcumin on TGF-β signaling in cancer cells and the role of TGF-β signaling in curcumin-induced anticancer activities have not been determined. Here, we investigate the role of curcumin on TGF-β signaling, and whether TGF-β signaling is involved in the antitumor activities of curcumin.
Methods: Human non-small cell lung cancer (NSCLC) cell lines, ACC-LC-176 (without TGF-β signaling), H358, and A549 (with TGF-β signaling) were treated with curcumin to determine cell growth, apoptosis, and tumorigenicity. Antitumor activities of curcumin were determined using these cell lines and an in vivo mouse model. We also tested the effect of curcumin on TGF-β/Smad signaling by western blotting and by luciferase assays.
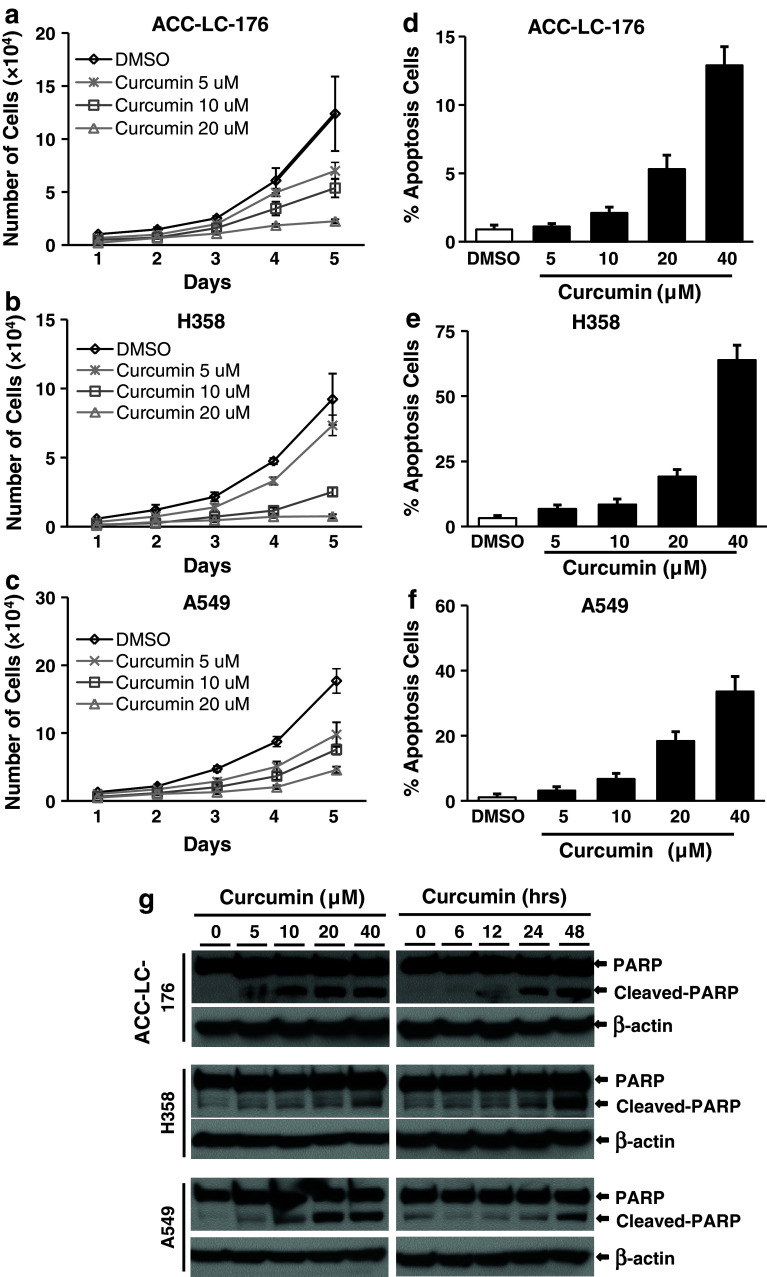
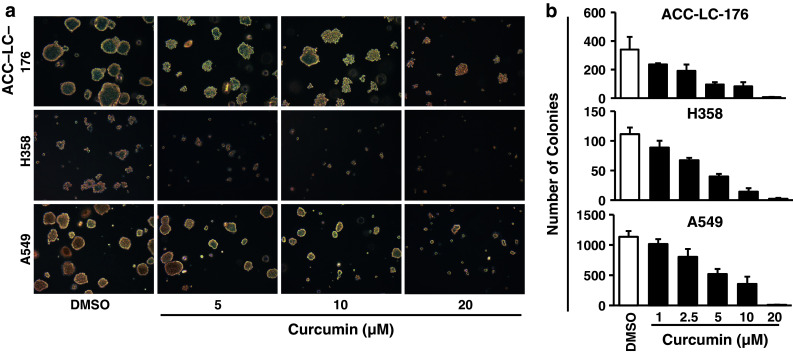
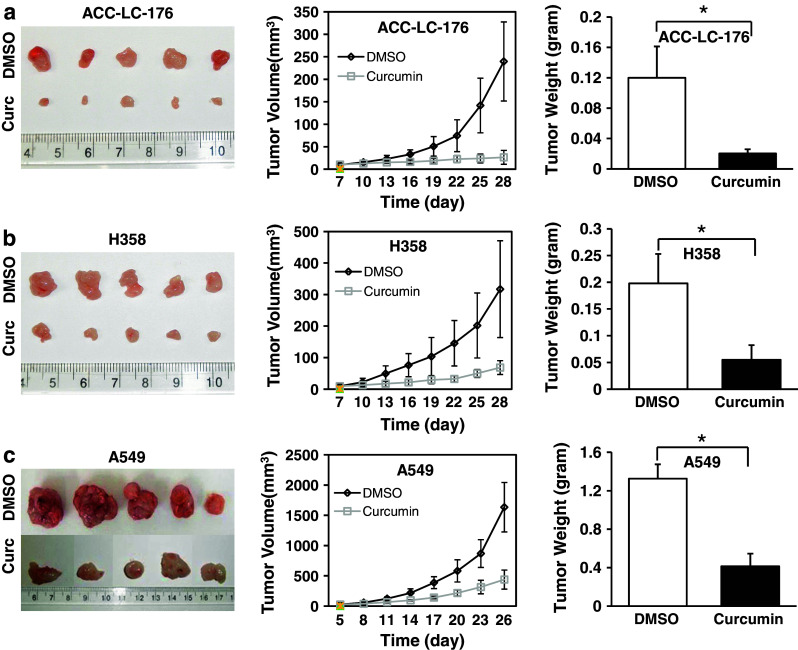
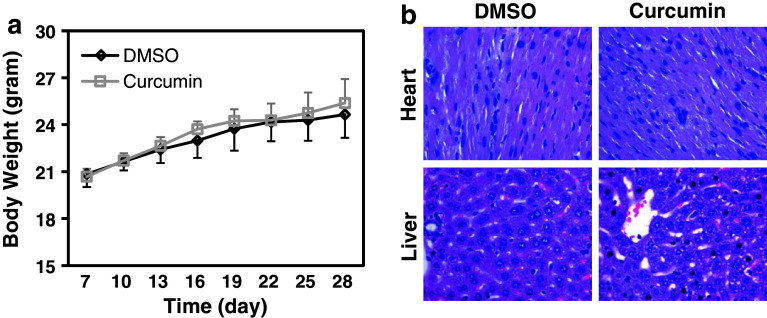
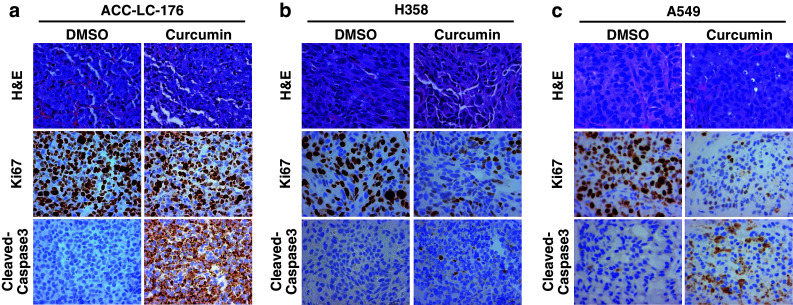
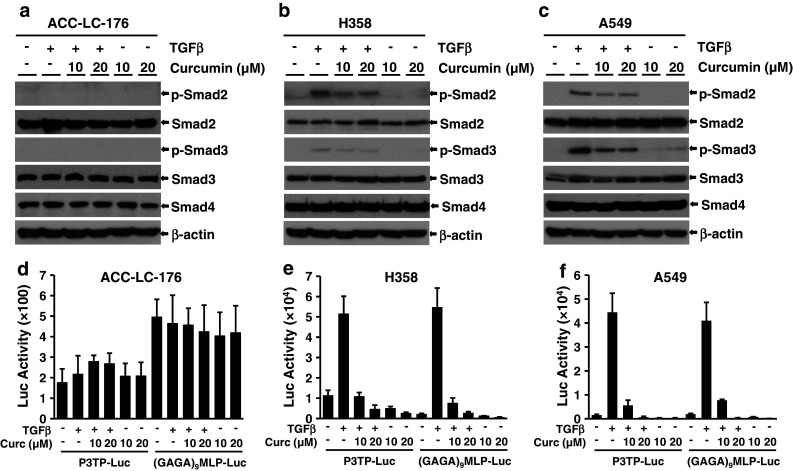
Results: Curcumin inhibited cell growth and induced apoptosis of all three NSCLC cell lines in vitro and in vivo. It significantly reduced subcutaneous tumor growth by these three cell lines irrespective of TGF-β signaling status. Curcumin inhibited TGF-β-induced Smad2/3 phosphorylation and transcription in H358 and A549 cells, but not in ACC-LC-176 cells.
Conclusions: Curcumin reduces tumorigenicity of human lung cancer cells in vitro and in vivo by inhibiting cell proliferation and promoting apoptosis. These results suggest that TGF-β signaling is not directly involved in curcumin-mediated growth inhibition, induction of apoptosis, and inhibition of tumorigenicity.
Conflict of interest statement
None.
Figures
Similar articles
-
Restoration of TGF-beta signalling reduces tumorigenicity in human lung cancer cells.Br J Cancer. 2005 Nov 14;93(10):1157-67. doi: 10.1038/sj.bjc.6602831. Br J Cancer. 2005. PMID: 16251876 Free PMC article.
-
Curcumin induces EGFR degradation in lung adenocarcinoma and modulates p38 activation in intestine: the versatile adjuvant for gefitinib therapy.PLoS One. 2011;6(8):e23756. doi: 10.1371/journal.pone.0023756. Epub 2011 Aug 17. PLoS One. 2011. PMID: 21858220 Free PMC article.
-
Effects of TGF-β signaling blockade on human A549 lung adenocarcinoma cell lines.Mol Med Rep. 2011 Sep-Oct;4(5):1007-15. doi: 10.3892/mmr.2011.530. Epub 2011 Jul 1. Mol Med Rep. 2011. PMID: 21725601
-
TGF-beta signaling and the role of inhibitory Smads in non-small cell lung cancer.J Thorac Oncol. 2010 Apr;5(4):417-9. doi: 10.1097/JTO.0b013e3181ce3afd. J Thorac Oncol. 2010. PMID: 20107423 Free PMC article. Review.
-
Molecular mechanism of TGF-β signaling pathway in colon carcinogenesis and status of curcumin as chemopreventive strategy.Tumour Biol. 2014 Aug;35(8):7295-305. doi: 10.1007/s13277-014-1840-1. Epub 2014 Mar 26. Tumour Biol. 2014. PMID: 24668546 Review.
Cited by
-
Targeting transforming growth factor-β signalling for cancer prevention and intervention: Recent advances in developing small molecules of natural origin.Clin Transl Med. 2022 Apr;12(4):e795. doi: 10.1002/ctm2.795. Clin Transl Med. 2022. PMID: 35384373 Free PMC article. Review.
-
Circulating Tumour Cells (CTCs) in NSCLC: From Prognosis to Therapy Design.Pharmaceutics. 2021 Nov 5;13(11):1879. doi: 10.3390/pharmaceutics13111879. Pharmaceutics. 2021. PMID: 34834295 Free PMC article. Review.
-
Curcumin as a potential inhibitor of TGFβ3 computational insights for breast cancer therapy.Sci Rep. 2025 Jan 22;15(1):2871. doi: 10.1038/s41598-025-86289-0. Sci Rep. 2025. PMID: 39843618 Free PMC article.
-
RNA-Seq and Network Analysis Revealed Interacting Pathways in TGF-β-Treated Lung Cancer Cell Lines.Cancer Inform. 2015 Apr 1;13(Suppl 5):129-40. doi: 10.4137/CIN.S14073. eCollection 2014. Cancer Inform. 2015. PMID: 25991908 Free PMC article.
-
Mechanistic insights curcumin's anti-inflammatory in pancreatic cancer: experimental and computational evidence implicating IL1B interference via IL10RA upregulation and NLRP3/TLR3 downregulation.Front Cell Dev Biol. 2025 Jun 4;13:1601908. doi: 10.3389/fcell.2025.1601908. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40535567 Free PMC article.
References
-
- Bayet-Robert M, Kwiatkowski F, Leheurteur M, Gachon F, Planchat E, Abrial C, Mouret-Reynier MA, Durando X, Barthomeuf C, Chollet P (2010) Phase I dose escalation trial of docetaxel plus curcumin in patients with advanced and metastatic breast cancer. Cancer Biol Ther 9(1):8–14 - PubMed
-
- Cheng AL, Hsu CH, Lin JK, Hsu MM, Ho YF, Shen TS, Ko JY, Lin JT, Lin BR, Ming-Shiang W, Yu HS, Jee SH, Chen GS, Chen TM, Chen CA, Lai MK, Pu YS, Pan MH, Wang YJ, Tsai CC, Hsieh CY (2001) Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res 21(4B):2895–2900 - PubMed
-
- Derynck R, Zhang YE (2003) Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 425(6958):577–584 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
