

DnaN clamp zones provide a platform for spatiotemporal coupling of mismatch detection to DNA replication
- PMID: 23228104
- PMCID: PMC5938748
- DOI: 10.1111/mmi.12115
DnaN clamp zones provide a platform for spatiotemporal coupling of mismatch detection to DNA replication
Abstract
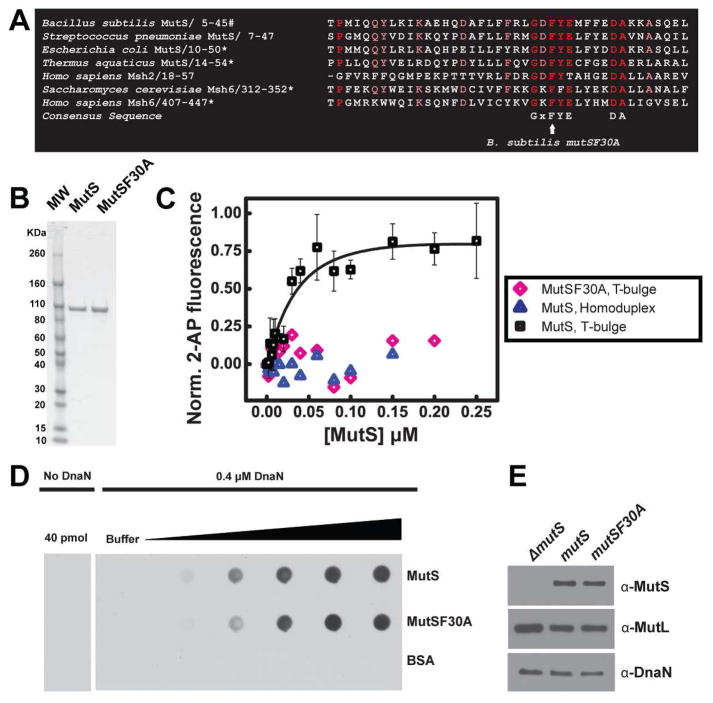
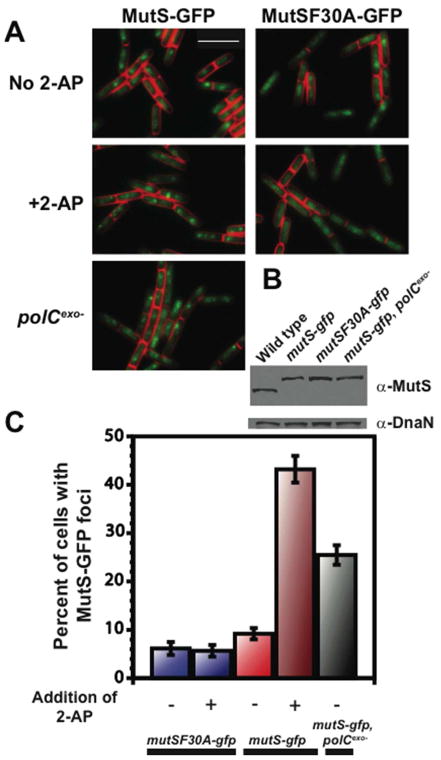
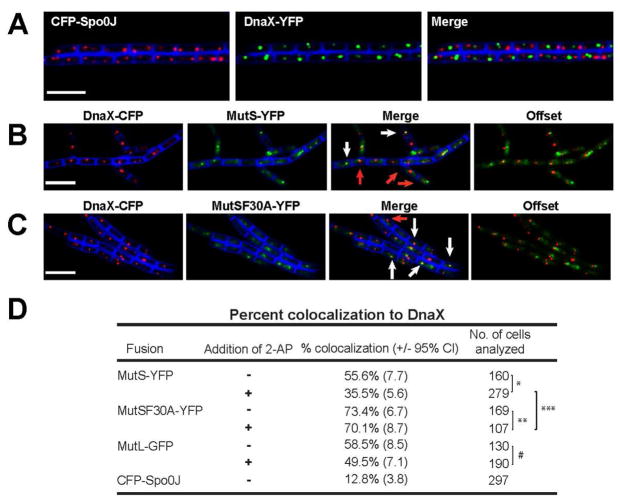
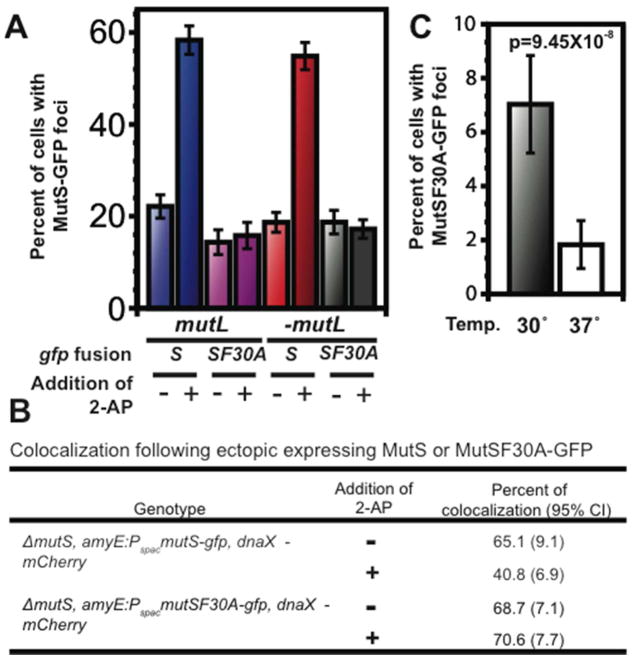
Mismatch repair (MMR) increases the fidelity of DNA replication by identifying and correcting replication errors. Processivity clamps are vital components of DNA replication and MMR, yet the mechanism and extent to which they participate in MMR remains unclear. We investigated the role of the Bacillus subtilis processivity clamp DnaN, and found that it serves as a platform for mismatch detection and coupling of repair to DNA replication. By visualizing functional MutS fluorescent fusions in vivo, we find that MutS forms foci independent of mismatch detection at sites of replication (i.e. the replisome). These MutS foci are directed to the replisome by DnaN clamp zones that aid mismatch detection by targeting the search to nascent DNA. Following mismatch detection, MutS disengages from the replisome, facilitating repair. We tested the functional importance of DnaN-mediated mismatch detection for MMR, and found that it accounts for 90% of repair. This high dependence on DnaN can be bypassed by increasing MutS concentration within the cell, indicating a secondary mode of detection in vivo whereby MutS directly finds mismatches without associating with the replisome. Overall, our results provide new insight into the mechanism by which DnaN couples mismatch recognition to DNA replication in living cells.
© 2012 Blackwell Publishing Ltd.
Figures
Similar articles
-
Single-molecule motions and interactions in live cells reveal target search dynamics in mismatch repair.Proc Natl Acad Sci U S A. 2015 Dec 15;112(50):E6898-906. doi: 10.1073/pnas.1507386112. Epub 2015 Nov 2. Proc Natl Acad Sci U S A. 2015. PMID: 26575623 Free PMC article.
-
Mismatch repair causes the dynamic release of an essential DNA polymerase from the replication fork.Mol Microbiol. 2011 Nov;82(3):648-63. doi: 10.1111/j.1365-2958.2011.07841.x. Epub 2011 Sep 30. Mol Microbiol. 2011. PMID: 21958350 Free PMC article.
-
Spatial coupling between DNA replication and mismatch repair in Caulobacter crescentus.Nucleic Acids Res. 2021 Apr 6;49(6):3308-3321. doi: 10.1093/nar/gkab112. Nucleic Acids Res. 2021. PMID: 33677508 Free PMC article.
-
Mismatch repair in Gram-positive bacteria.Res Microbiol. 2016 Jan;167(1):4-12. doi: 10.1016/j.resmic.2015.08.006. Epub 2015 Sep 3. Res Microbiol. 2016. PMID: 26343983 Review.
-
Single-molecule views of MutS on mismatched DNA.DNA Repair (Amst). 2014 Aug;20:82-93. doi: 10.1016/j.dnarep.2014.02.014. Epub 2014 Mar 12. DNA Repair (Amst). 2014. PMID: 24629484 Free PMC article. Review.
Cited by
-
Using stable MutS dimers and tetramers to quantitatively analyze DNA mismatch recognition and sliding clamp formation.Nucleic Acids Res. 2013 Sep;41(17):8166-81. doi: 10.1093/nar/gkt582. Epub 2013 Jul 1. Nucleic Acids Res. 2013. PMID: 23821665 Free PMC article.
-
DNA replication initiation timing is important for maintaining genome integrity.bioRxiv [Preprint]. 2024 Jun 18:2024.06.18.599555. doi: 10.1101/2024.06.18.599555. bioRxiv. 2024. Update in: J Bacteriol. 2025 Aug 21;207(8):e0017525. doi: 10.1128/jb.00175-25. PMID: 38948856 Free PMC article. Updated. Preprint.
-
A bacterial toxin inhibits DNA replication elongation through a direct interaction with the β sliding clamp.Mol Cell. 2013 Dec 12;52(5):617-28. doi: 10.1016/j.molcel.2013.10.014. Epub 2013 Nov 14. Mol Cell. 2013. PMID: 24239291 Free PMC article.
-
Mismatch binding, ADP-ATP exchange and intramolecular signaling during mismatch repair.DNA Repair (Amst). 2016 Feb;38:24-31. doi: 10.1016/j.dnarep.2015.11.017. Epub 2015 Dec 2. DNA Repair (Amst). 2016. PMID: 26704427 Free PMC article. Review.
-
MutS regulates access of the error-prone DNA polymerase Pol IV to replication sites: a novel mechanism for maintaining replication fidelity.Nucleic Acids Res. 2016 Sep 19;44(16):7700-13. doi: 10.1093/nar/gkw494. Epub 2016 Jun 1. Nucleic Acids Res. 2016. PMID: 27257069 Free PMC article.
References
-
- Alani E, Chi NW, Kolodner R. The Saccharomyces cerevisiae Msh2 protein specifically binds to duplex oligonucleotides containing mismatched DNA base pairs and insertions. Genes & development. 1995;9:234–247. - PubMed
-
- Berkmen MB, Grossman AD. Spatial and temporal organization of the Bacillus subtilis replication cycle. Mol Microbiol. 2006;62:57–71. - PubMed
-
- Bowers J, Tran PT, Liskay RM, Alani E. Analysis of yeast MSH2-MSH6 suggests that the initiation of mismatch repair can be separated into discrete steps. J Mol Biol. 2000;302:327–338. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
