

Impairment of cortical GABAergic synaptic transmission in an environmental rat model of autism
- PMID: 23228615
- PMCID: PMC3674140
- DOI: 10.1017/S1461145712001216
Impairment of cortical GABAergic synaptic transmission in an environmental rat model of autism
Abstract
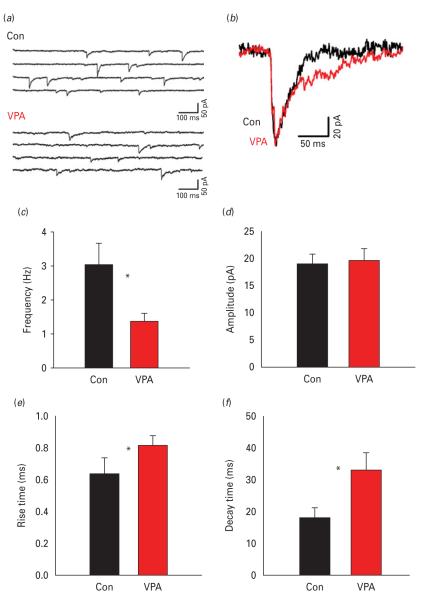
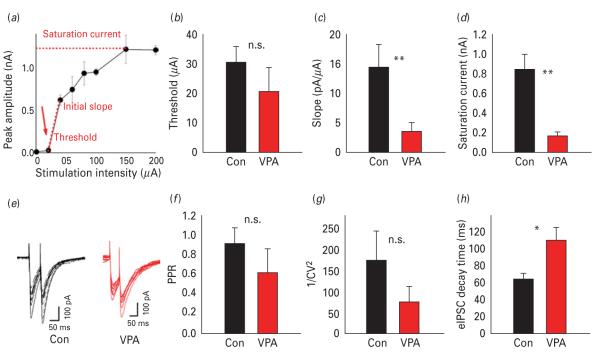
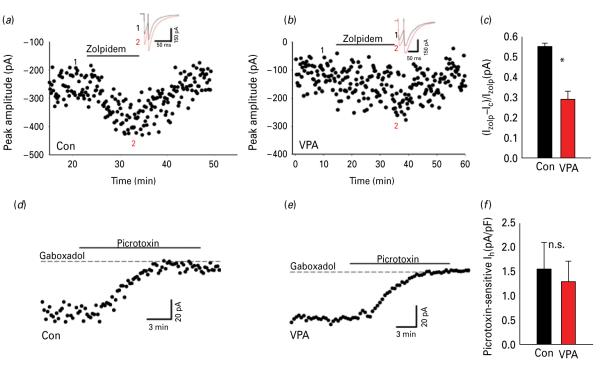
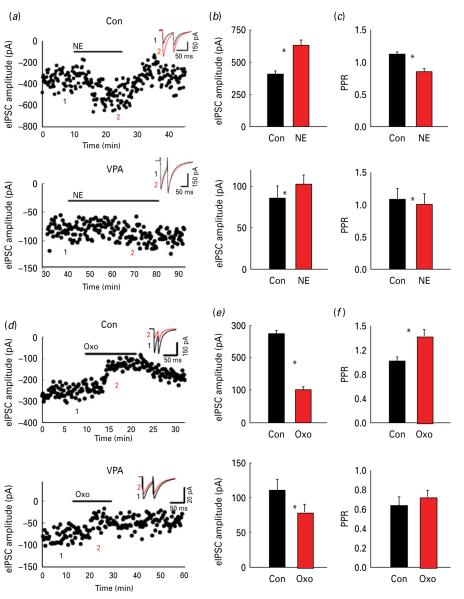
The biological mechanisms of autism spectrum disorders (ASDs) are largely unknown in spite of extensive research. ASD is characterized by altered function of multiple brain areas including the temporal cortex and by an increased synaptic excitation:inhibition ratio. While numerous studies searched for evidence of increased excitation in ASD, fewer have investigated the possibility of reduced inhibition. We characterized the cortical γ-amino butyric acid (GABA)ergic system in the rat temporal cortex of an ASD model [offspring of mothers prenatally injected with valproic acid (VPA)], by monitoring inhibitory post-synaptic currents (IPSCs) with patch-clamp. We found that numerous features of inhibition were severely altered in VPA animals compared to controls. Among them were the frequency of miniature IPSCs, the rise time and decay time of electrically-evoked IPSCs, the slope and saturation of their input/output curves, as well as their modulation by adrenergic and muscarinic agonists and by the synaptic GABAA receptor allosteric modulator zolpidem (but not by the extra-synaptic modulator gaboxadol). Our data suggest that both pre- and post-synaptic, but not extra-synaptic, inhibitory transmission is impaired in the offspring of VPA-injected mothers. We speculate that impairment in the GABAergic system critically contributes to an increase in the ratio between synaptic excitation and inhibition, which in genetically predisposed individuals may alter cortical circuits responsible for emotional, communication and social impairments at the core of ASD.
Figures
References
-
- Antshel KM, Polacek C, McMahon M, Dygert K, Spenceley L, Dygert L, Miller L, Faisal F. Comorbid ADHD and anxiety affect social skills group intervention treatment efficacy in children with autism spectrum disorders. J Dev Behav Pediatr. 2011;32:439–446. - PubMed
-
- Atzori M, Lei S, Evans DI, Kanold PO, Phillips-Tansey E, McIntyre O, McBain CJ. Differential synaptic processing separates stationary from transient inputs to the auditory cortex. Nat Neurosci. 2001;4:1230–1237. - PubMed
-
- Blatt GJ, Fitzgerald CM, Guptill JT, Booker AB, Kemper TL, Bauman ML. Density and distribution of hippocampal neurotransmitter receptors in autism: an autoradiographic study. J Autism Dev Disord. 2001;31:537–543. - PubMed
-
- Bourgeron T. The possible interplay of synaptic and clock genes in autism spectrum disorders. Cold Spring Harb Symp Quant Biol. 2007;72:645–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
