

Review
doi: 10.1083/jcb.201210040.
Linking abnormal mitosis to the acquisition of DNA damage
Affiliations
- PMID: 23229895
- PMCID: PMC3518222
- DOI: 10.1083/jcb.201210040
Item in Clipboard
Review
Linking abnormal mitosis to the acquisition of DNA damage
J Cell Biol.
.
Abstract
Cellular defects that impair the fidelity of mitosis promote chromosome missegregation and aneuploidy. Increasing evidence reveals that errors in mitosis can also promote the direct and indirect acquisition of DNA damage and chromosome breaks. Consequently, deregulated cell division can devastate the integrity of the normal genome and unleash a variety of oncogenic stimuli that may promote transformation. Recent work has shed light on the mechanisms that link abnormal mitosis with the development of DNA damage, how cells respond to such affronts, and the potential impact on tumorigenesis.
Figures
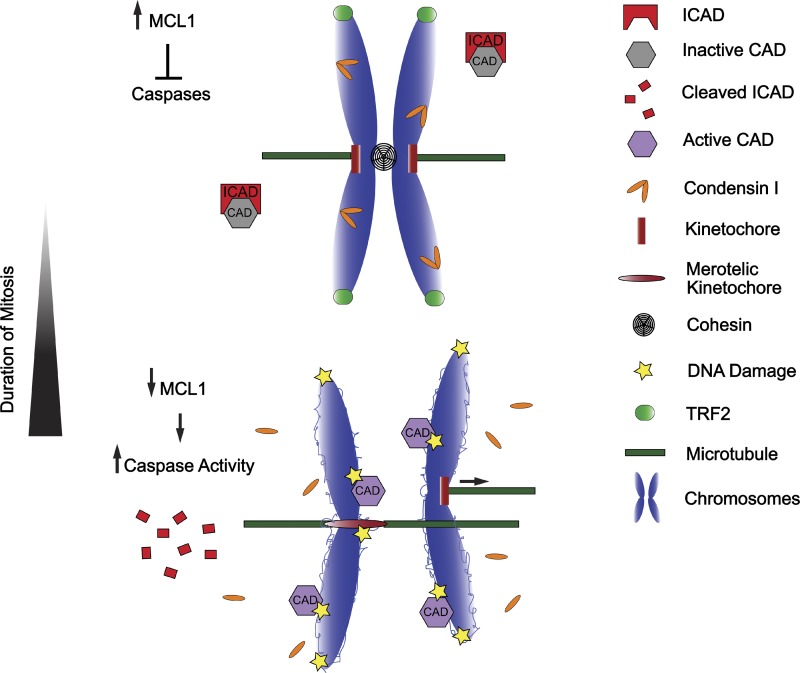
Prolonged mitosis gives rise to DNA damage through multiple mechanisms. Prolonged mitosis leads to depletion of many proteins, including anti-apoptotic proteins such as MCL1. This induces a partial caspase activation and destruction of ICAD and CAP-H, which frees the DNase CAD to cleave decondensed loops on chromosomal DNA. In addition, the telomere-protecting protein TRF2 loses its telomeric localization, exposing the linear ends of chromosomes that are recognized as DSBs. Finally, sister chromatid cohesion is gradually lost during prolonged mitosis, and this “cohesion fatigue” promotes premature sister chromatid separation and merotelic attachment. The forces generated by merotelic attachment at kinetochores may combine with preexisting DNA damage at the underlying centromeres to generate arm-level chromosome breaks, an anomaly commonly observed in human cancers.
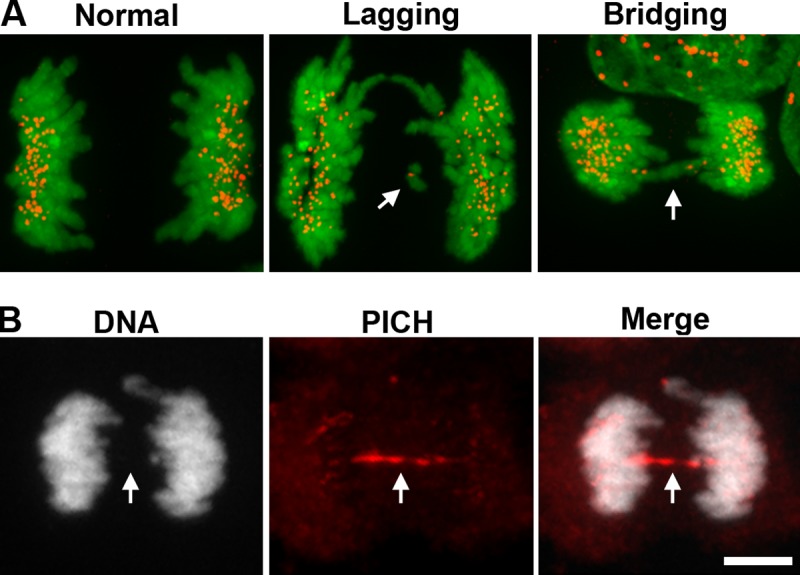
Anaphase lagging chromosomes versus chromosome bridges. (A) Anaphase lagging chromosomes (“Lagging”, white arrow) are identified as single, kinetochore-positive chromosomes that lag between the two masses of segregating chromosomes during anaphase. Lagging chromosomes are commonly caused by merotelic attachments. By contrast, pathological chromosome bridges (“Bridging”, white arrow) completely span the segregating masses of chromosomes during anaphase. Pathological bridges are caused by condensation and cohesion defects, or by dicentric chromosomes being pulled to opposite poles, and are readily visualized with DNA-intercalating dyes (kinetochores, red; chromosomes stained with the DNA intercalating dye Hoechst, green). Images reproduced from Ganem et al. (2009). (B) Ultra-fine chromosome bridges (white arrow) result from inefficient decatenation of sister chromosomes and cannot be detected using DNA dyes, requiring instead detection with specific protein markers (PICH, red; chromosomes stained with the DNA intercalating dye Hoechst, white). Bar, 10 µm. Images courtesy of Taruho Kuroda (Harvard Medical School, Boston, MA).
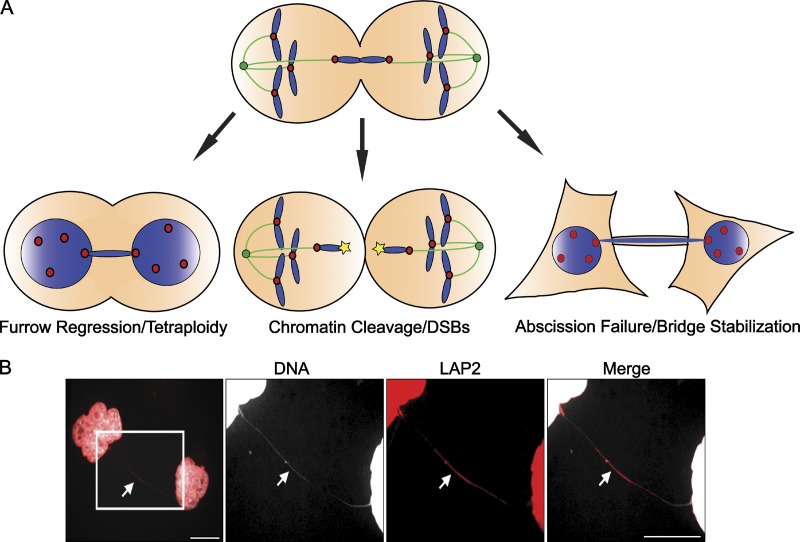
Consequences of chromosome bridges. (A) Chromosome bridges can promote cytokinetic furrow regression, chromosomal cleavage, or abscission failure. However, the factors governing which of these outcomes will occur remain entirely unknown. One possibility is that previously nicked or otherwise damaged chromosomes may be more susceptible to cleavage, whereas undamaged whole chromosomes may be more likely to promote furrow regression or abscission delays. (B) Chromosome bridges (white arrow) that are not cleaved or resolved during cytokinesis persist well into the following cell cycle, even stretching extensively as cells move apart (DNA, white; LAP2, red). Bars, 10 µm. Images courtesy of Taruho Kuroda (Harvard Medical School, Boston, MA).
References
-
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. 2002. Molecular Biology of the Cell. 4th ed Garland Science, NY: 1548 pp
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
