

Variant repeats are interspersed throughout the telomeres and recruit nuclear receptors in ALT cells
- PMID: 23229897
- PMCID: PMC3518223
- DOI: 10.1083/jcb.201207189
Variant repeats are interspersed throughout the telomeres and recruit nuclear receptors in ALT cells
Abstract
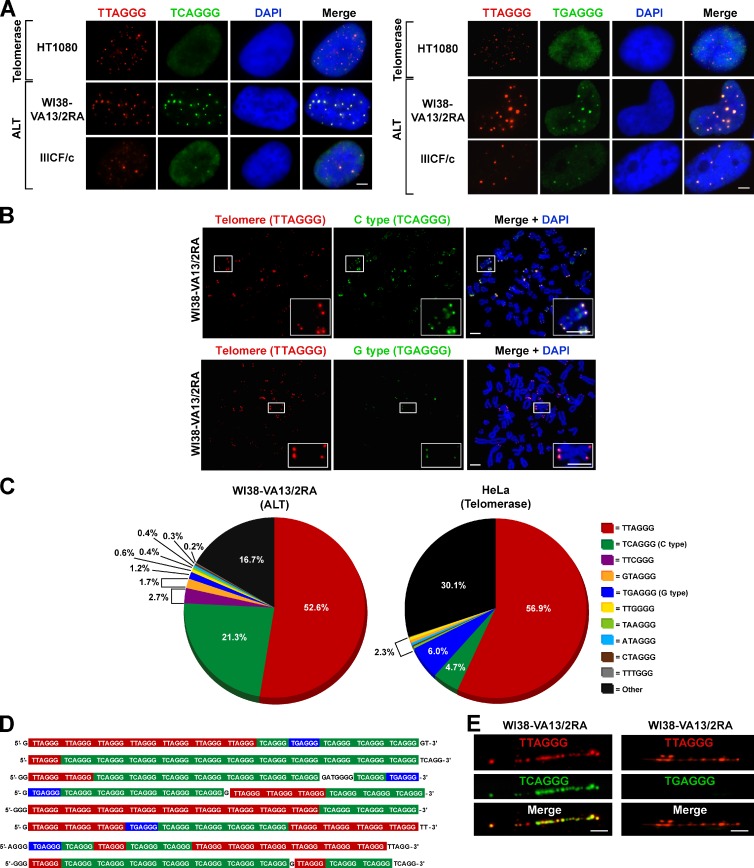
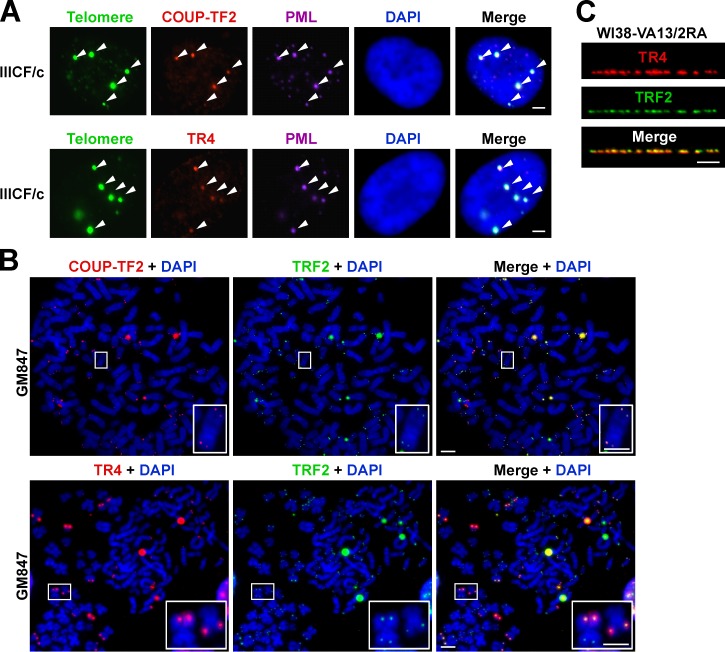
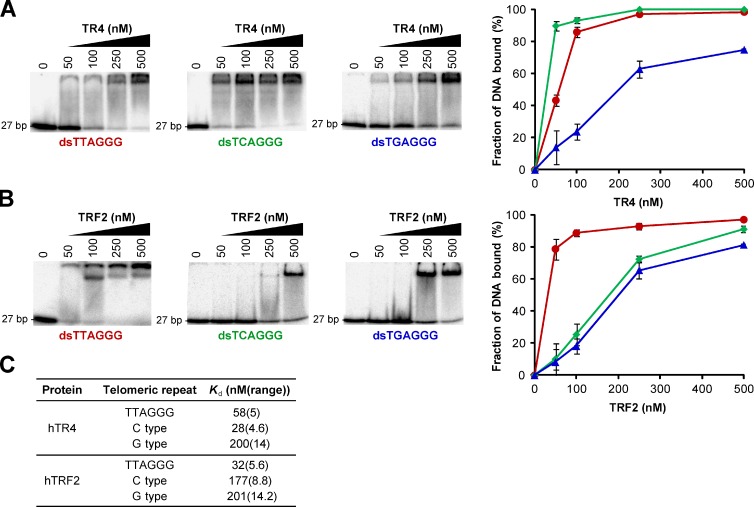
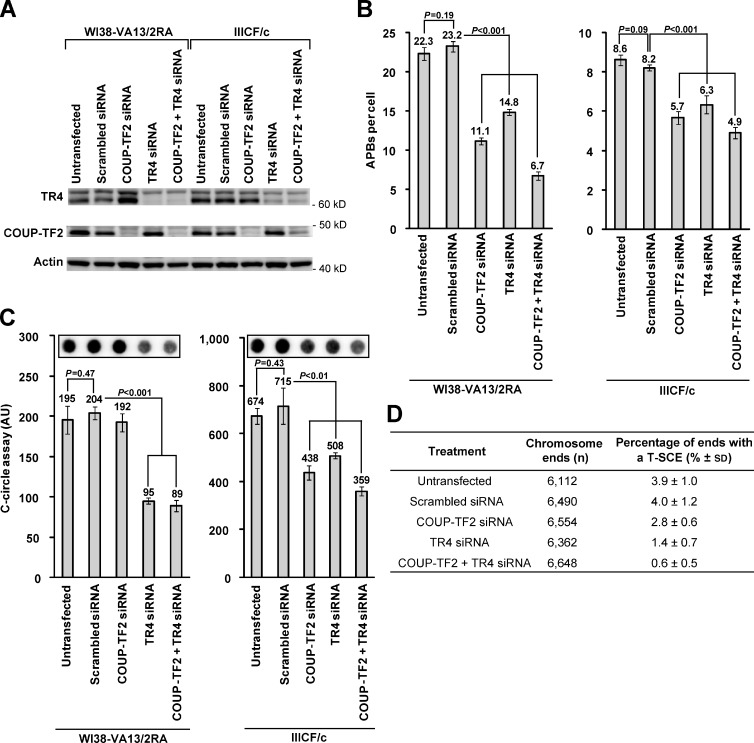
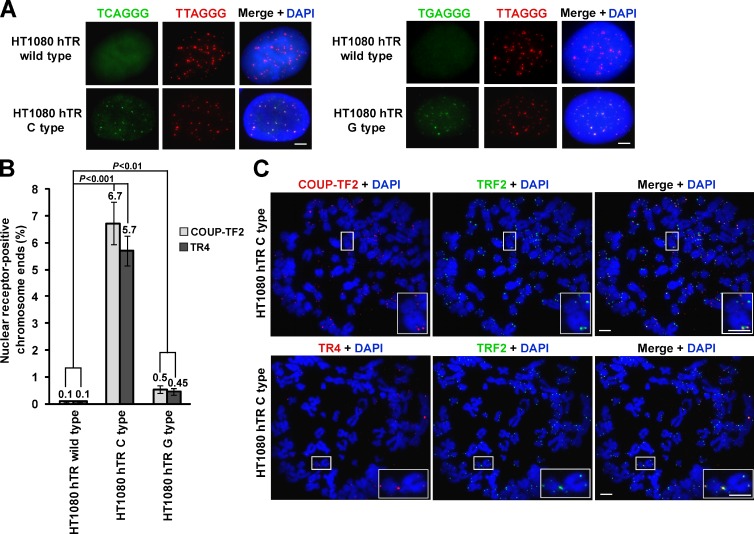
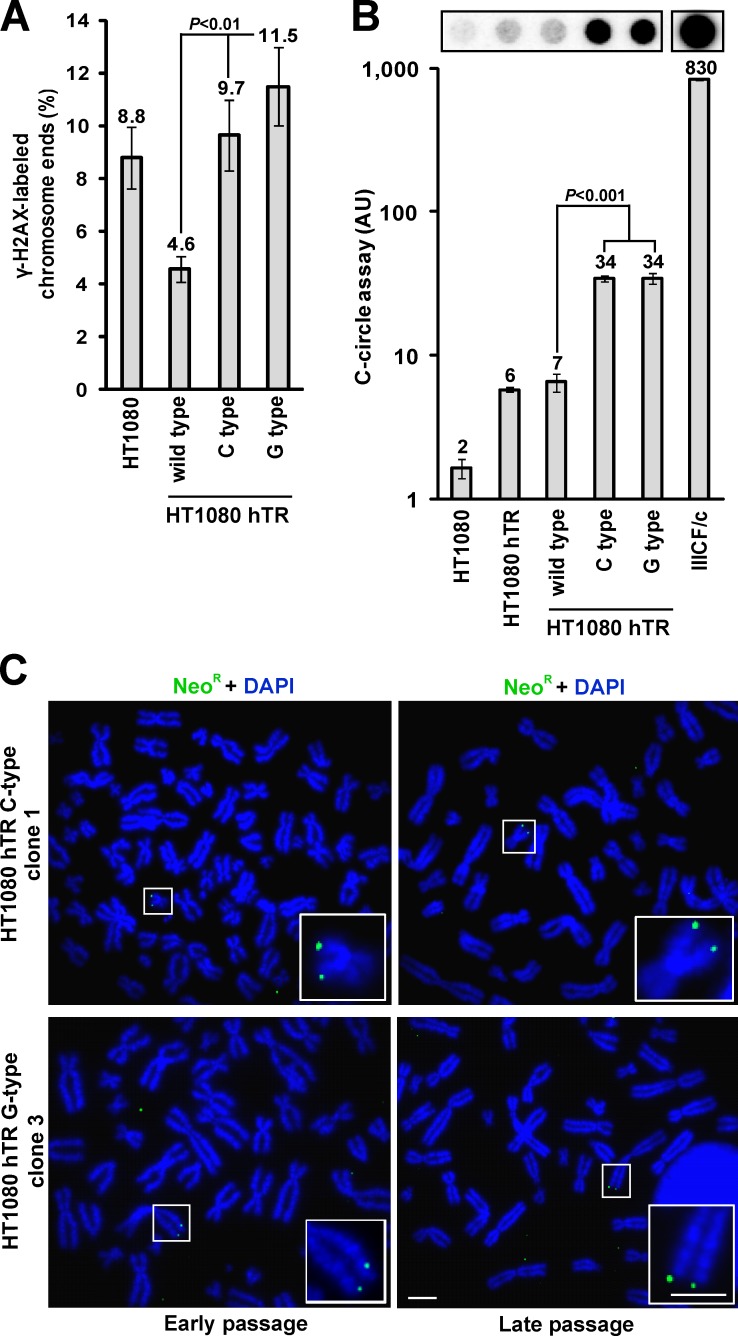
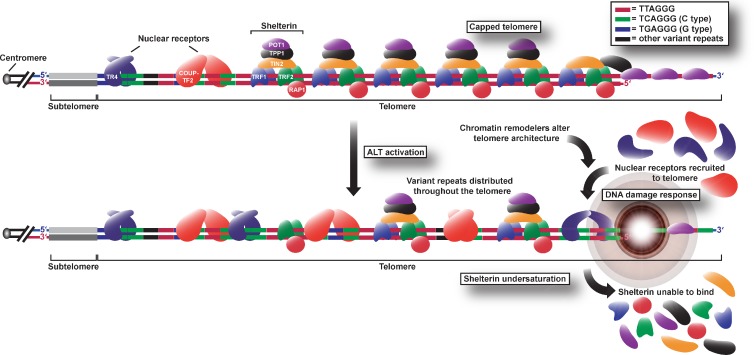
Telomeres in cells that use the recombination-mediated alternative lengthening of telomeres (ALT) pathway elicit a DNA damage response that is partly independent of telomere length. We therefore investigated whether ALT telomeres contain structural abnormalities that contribute to ALT activity. Here we used next generation sequencing to analyze the DNA content of ALT telomeres. We discovered that variant repeats were interspersed throughout the telomeres of ALT cells. We found that the C-type (TCAGGG) variant repeat predominated and created a high-affinity binding site for the nuclear receptors COUP-TF2 and TR4. Nuclear receptors were directly recruited to telomeres and ALT-associated characteristics were induced after incorporation of the C-type variant repeat by a mutant telomerase. We propose that the presence of variant repeats throughout ALT telomeres results from recombination-mediated telomere replication and spreading of variant repeats from the proximal regions of the telomeres and that the consequent binding of nuclear receptors alters the architecture of telomeres to facilitate further recombination.
Figures
References
-
- Bechard L.H., Butuner B.D., Peterson G.J., McRae W., Topcu Z., McEachern M.J. 2009. Mutant telomeric repeats in yeast can disrupt the negative regulation of recombination-mediated telomere maintenance and create an alternative lengthening of telomeres-like phenotype. Mol. Cell. Biol. 29:626–639 10.1128/MCB.00423-08 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
