

A high-throughput assay for phosphoprotein-specific phosphatase activity in cellular extracts
- PMID: 23233447
- PMCID: PMC3591670
- DOI: 10.1074/mcp.O112.024059
A high-throughput assay for phosphoprotein-specific phosphatase activity in cellular extracts
Abstract
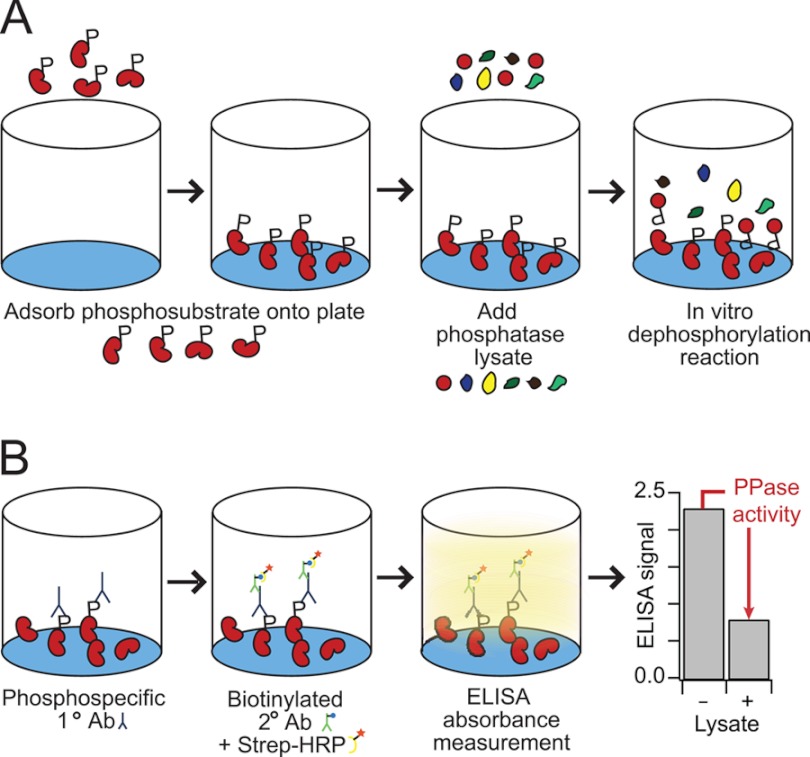
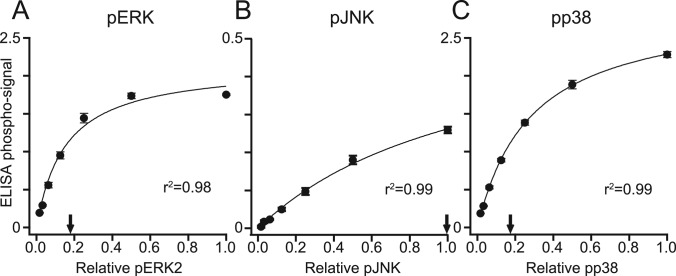
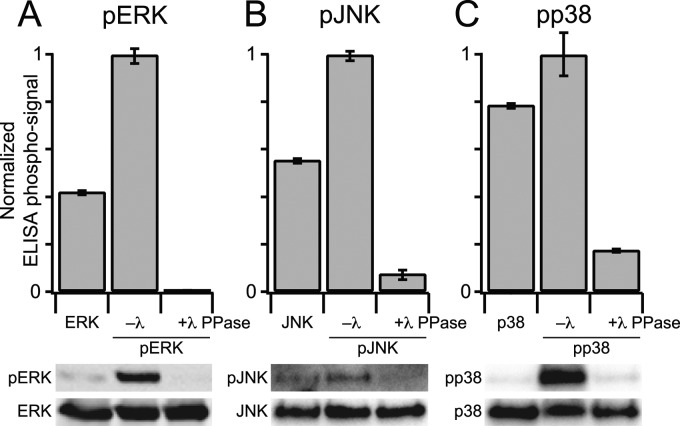
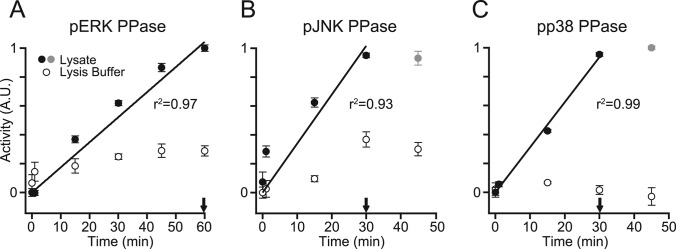
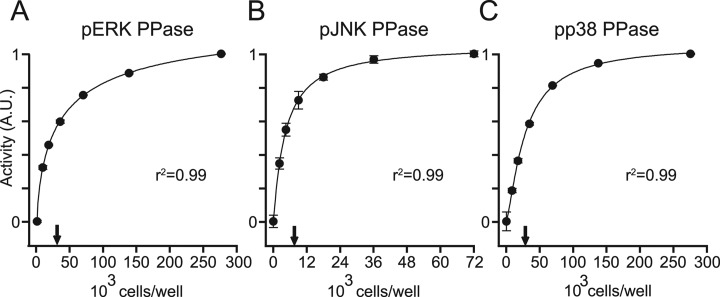
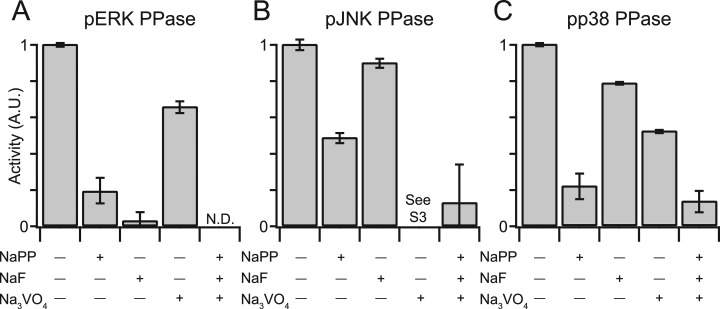
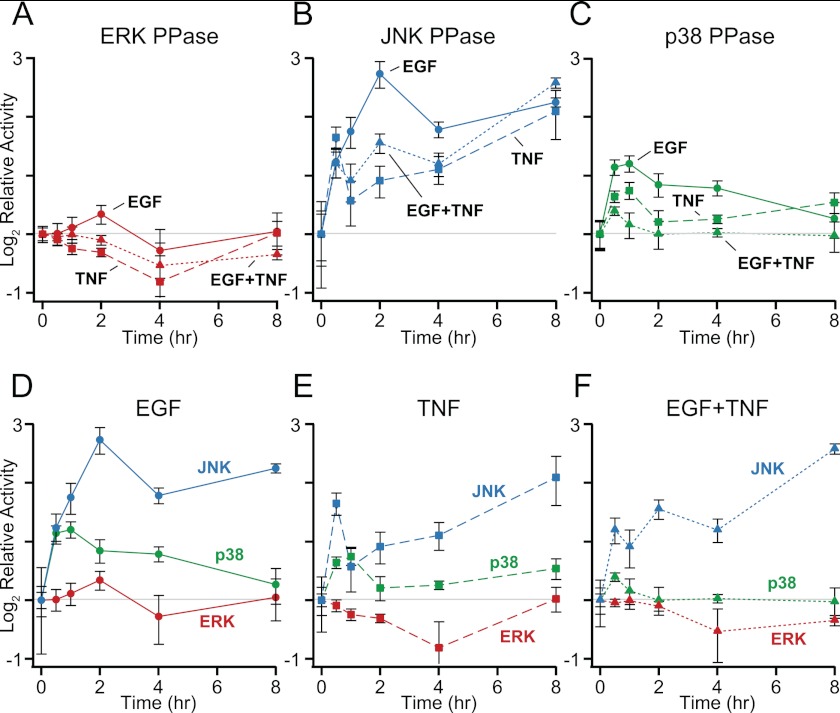
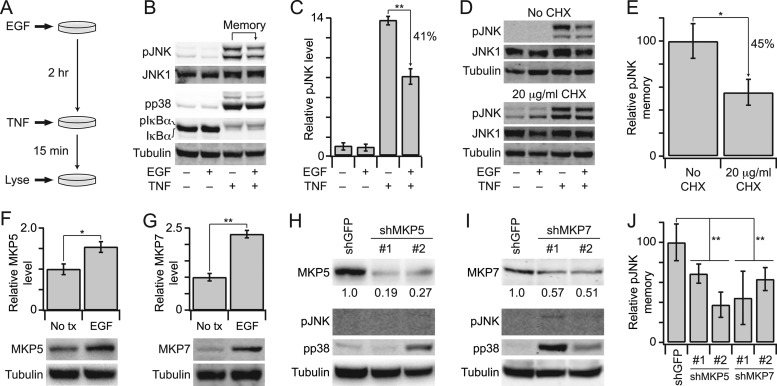
Protein phosphatases undo the post-translational modifications of kinase-signaling networks, but phosphatase activation in cells is difficult to measure and interpret. Here, we report the design of a quantitative and high-throughput assay platform for monitoring cellular phosphatase activity toward specific phosphoprotein targets. Protein substrates of interest are purified recombinantly, phosphorylated in vitro using the upstream kinase, and adsorbed to 96-well plates. Total phosphatase extracts from cells are then added to trigger a solid-phase dephosphorylation reaction. After stopping the reaction, phosphoprotein levels are quantified by ELISA with a phospho-specific antibody, and the loss of phospho-specific immunoreactivity is used as the readout of phosphatase activity. We illustrate the generality of the method by developing specific phosphatase-activity assays for the three canonical mitogen-activated protein phospho-kinases: ERK, JNK, and p38. The assays capture changes in activity with a dynamic range of 25-100-fold and are sensitive to a limit of detection below 25,000 cells. When applied to cytokine-induced signaling, the assays revealed complex and dynamic regulation of phosphatases suggesting cross-communication and a means for cellular memory. Our assay platform should be beneficial for phosphoproteomic surveys and computational-systems models of signaling, where phosphatases are known to be important but their activities are rarely measured.
Figures
References
-
- Tonks N. K. (2006) Protein tyrosine phosphatases: from genes, to function, to disease. Nat. Rev. Mol. Cell Biol. 7, 833–846 - PubMed
-
- Ingebritsen T. S., Cohen P. (1983) Protein phosphatases: properties and role in cellular regulation. Science 221, 331–338 - PubMed
-
- Keyse S. M. (2008) Dual-specificity MAP kinase phosphatases (MKPs) and cancer. Cancer Metastasis Rev. 27, 253–261 - PubMed
-
- Roy J., Cyert M. S. (2009) Cracking the phosphatase code: docking interactions determine substrate specificity. Science signaling 2, re9. - PubMed
-
- Shi Y. (2009) Serine/threonine phosphatases: mechanism through structure. Cell 139, 468–484 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
