

RapA2 is a calcium-binding lectin composed of two highly conserved cadherin-like domains that specifically recognize Rhizobium leguminosarum acidic exopolysaccharides
- PMID: 23235153
- PMCID: PMC3554953
- DOI: 10.1074/jbc.M112.411769
RapA2 is a calcium-binding lectin composed of two highly conserved cadherin-like domains that specifically recognize Rhizobium leguminosarum acidic exopolysaccharides
Abstract
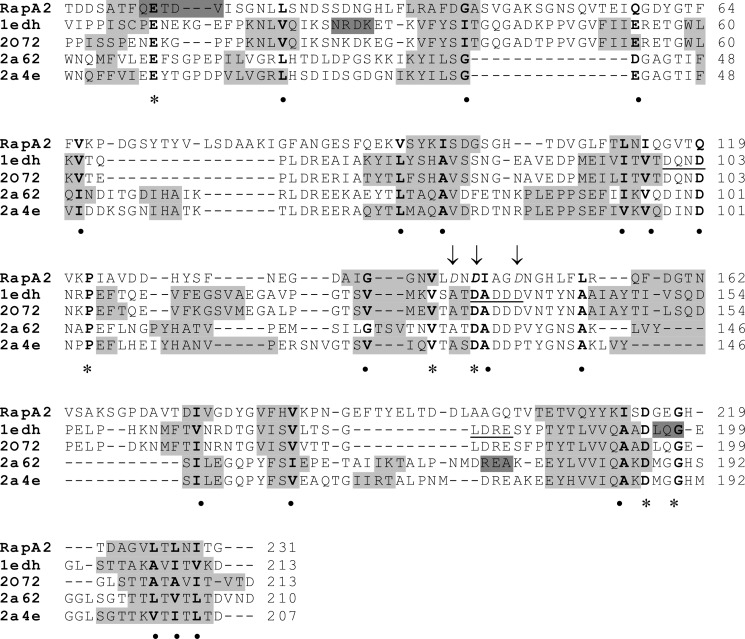
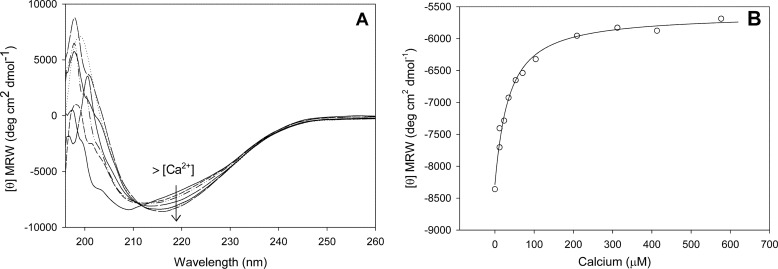
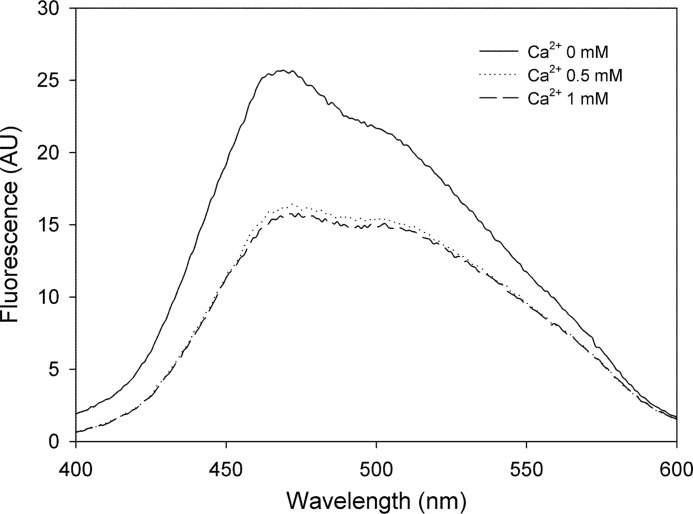
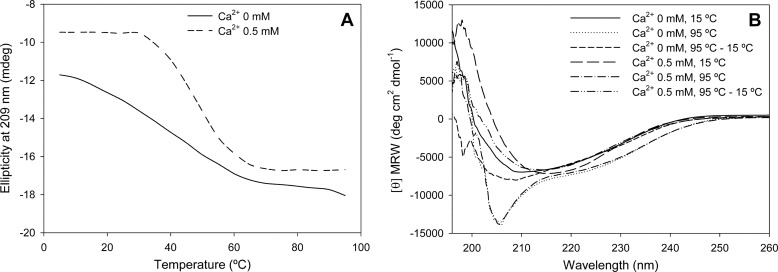
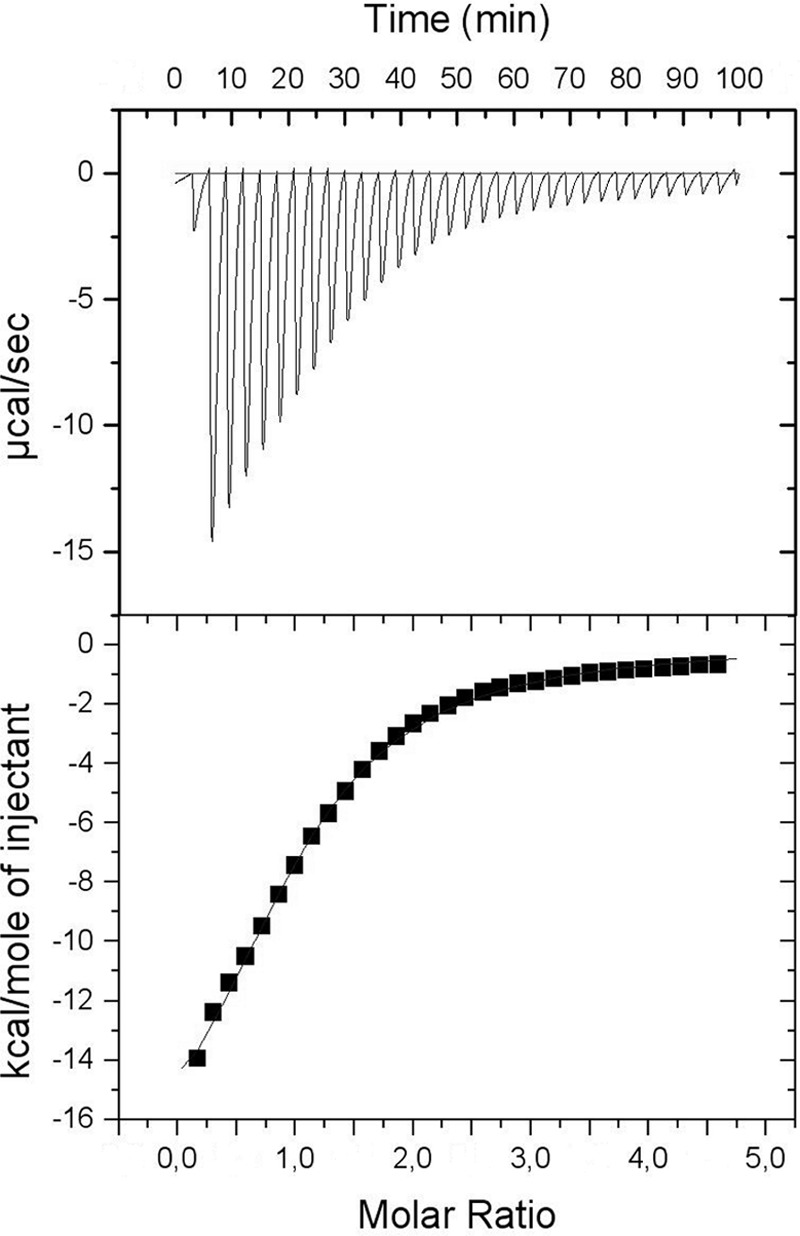
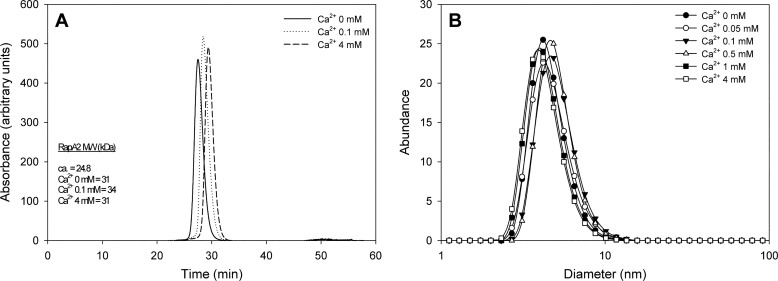
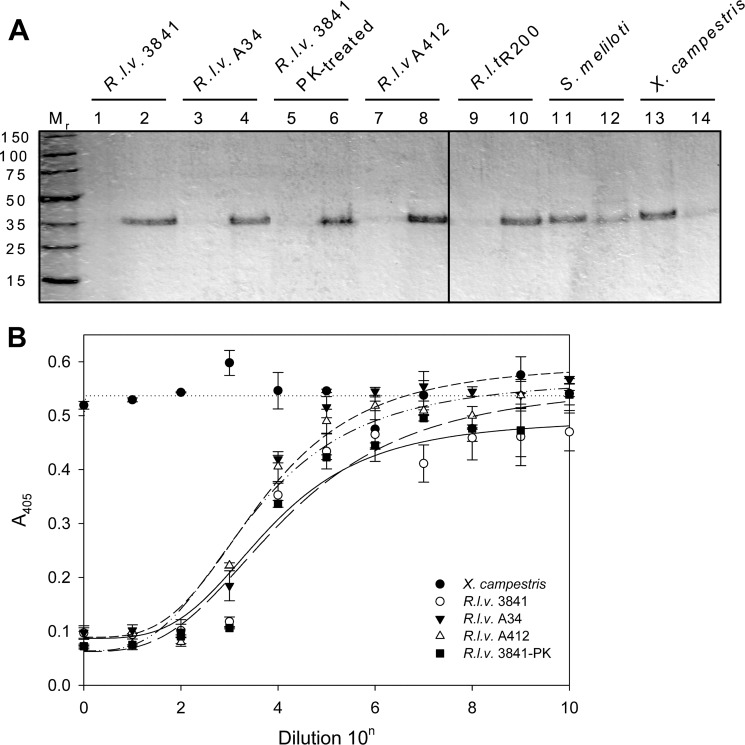
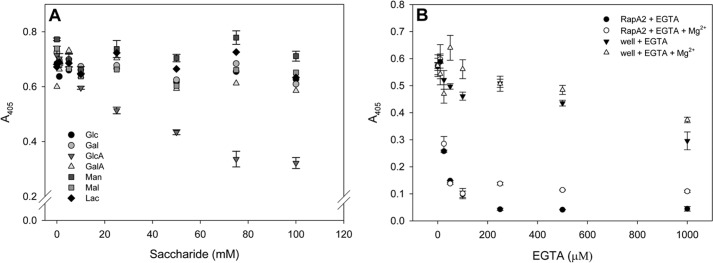
In silico analyses have revealed a conserved protein domain (CHDL) widely present in bacteria that has significant structural similarity to eukaryotic cadherins. A CHDL domain was shown to be present in RapA, a protein that is involved in autoaggregation of Rhizobium cells, biofilm formation, and adhesion to plant roots as shown by us and others. Structural similarity to cadherins suggested calcium-dependent oligomerization of CHDL domains as a mechanistic basis for RapA action. Here we show by circular dichroism spectroscopy, light scattering, isothermal titration calorimetry, and other methods that RapA2 from Rhizobium leguminosarum indeed exhibits a cadherin-like β-sheet conformation and that its proper folding and stability are dependent on the binding of one calcium ion per protein molecule. By further in silico analysis we also reveal that RapA2 consists of two CHDL domains and expand the range of CHDL-containing proteins in bacteria and archaea. However, light scattering assays at various concentrations of added calcium revealed that RapA2 formed neither homo-oligomers nor hetero-oligomers with RapB (a distinct CHDL protein), indicating that RapA2 does not mediate cellular interactions through a cadherin-like mechanism. Instead, we demonstrate that RapA2 interacts specifically with the acidic exopolysaccharides (EPSs) produced by R. leguminosarum in a calcium-dependent manner, sustaining a role of these proteins in the development of the biofilm matrix made of EPS. Because EPS binding by RapA2 can only be attributed to its two CHDL domains, we propose that RapA2 is a calcium-dependent lectin and that CHDL domains in various bacterial and archaeal proteins confer carbohydrate binding activity to these proteins.
Figures
References
-
- Downie J. A. (2010) The roles of extracellular proteins, polysaccharides, and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 34, 150–170 - PubMed
-
- Rinaudi L. V., Giordano W. (2010) An integrated view of biofilm formation in rhizobia. FEMS Microbiol. Lett. 304, 1–11 - PubMed
-
- Russo D. M., Williams A., Edwards A., Posadas D. M., Finnie C., Dankert M., Downie J. A., Zorreguieta A. (2006) Proteins exported via the PrsD-PrsE type I secretion system and the acidic exopolysaccharide are involved in biofilm formation by Rhizobium leguminosarum. J. Bacteriol. 188, 4474–4486 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
