

Purification and characterization of Escherichia coli MreB protein
- PMID: 23235161
- PMCID: PMC3561565
- DOI: 10.1074/jbc.M112.413708
Purification and characterization of Escherichia coli MreB protein
Abstract
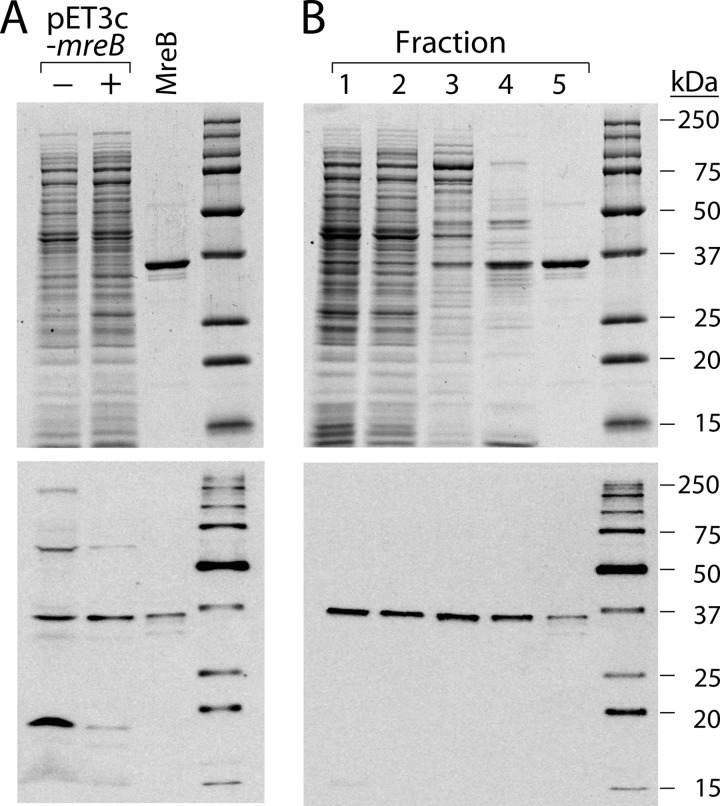
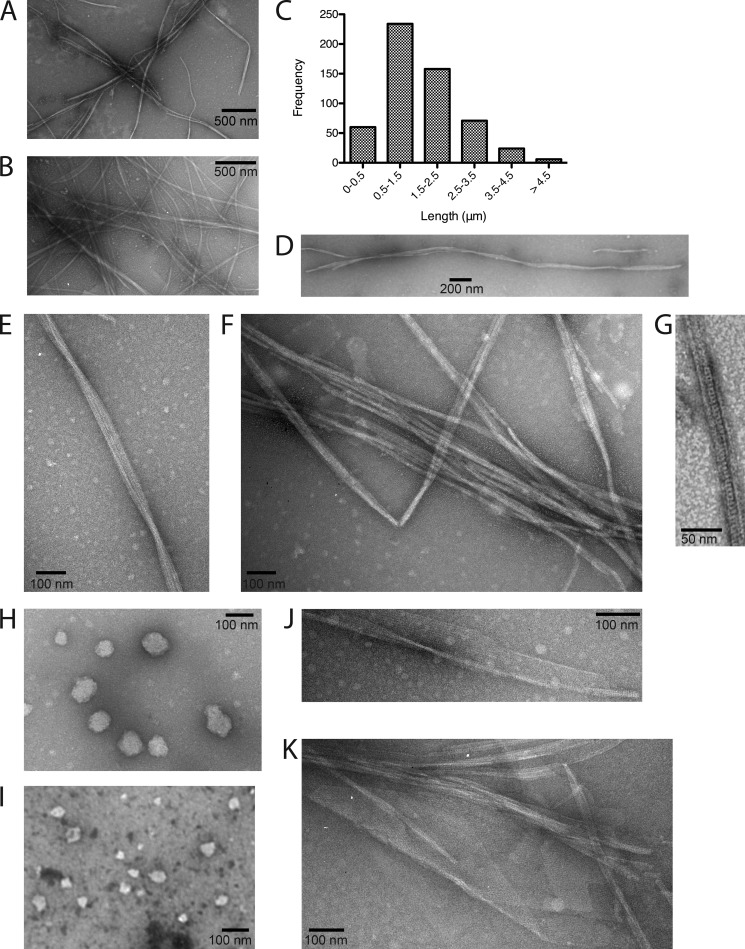
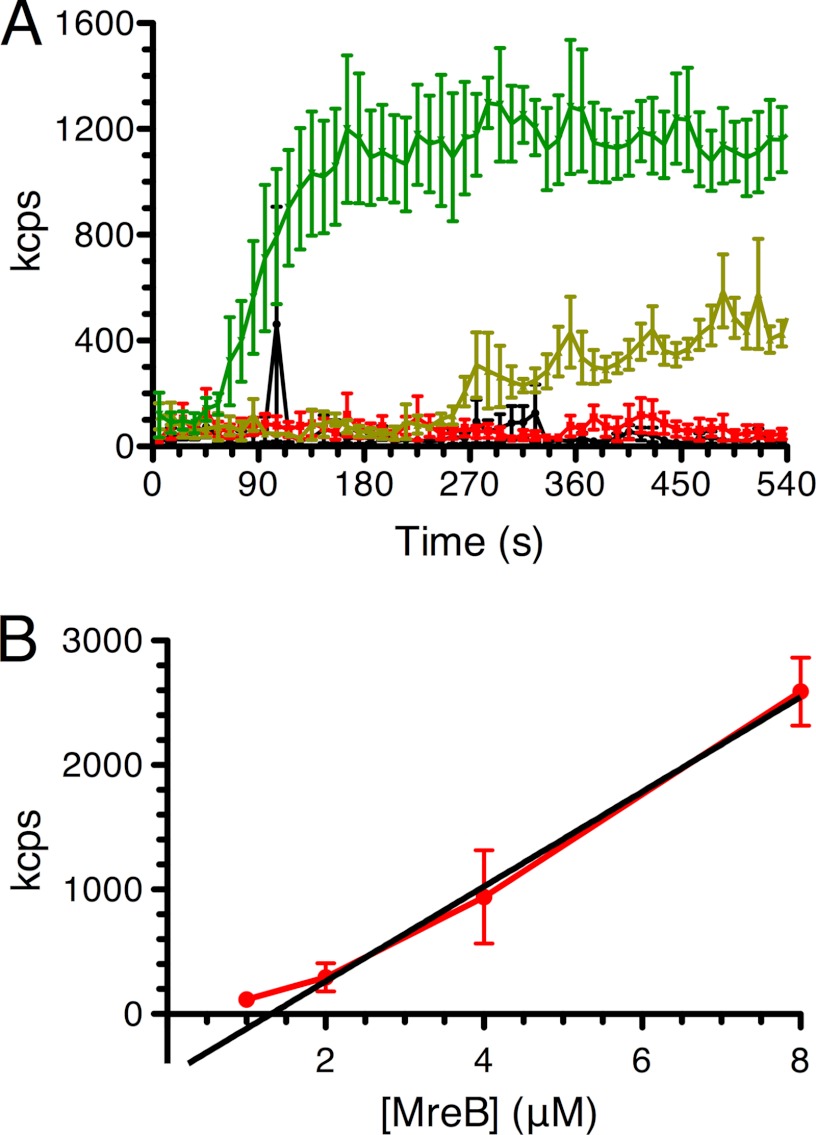
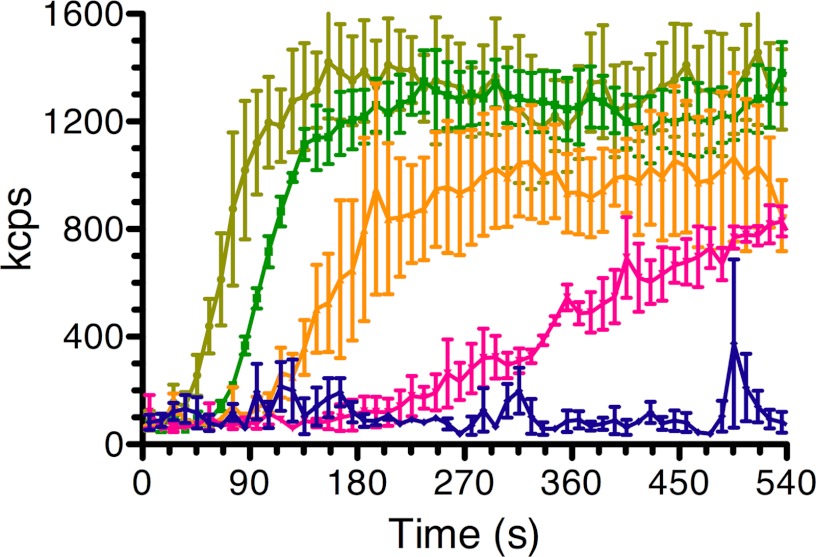
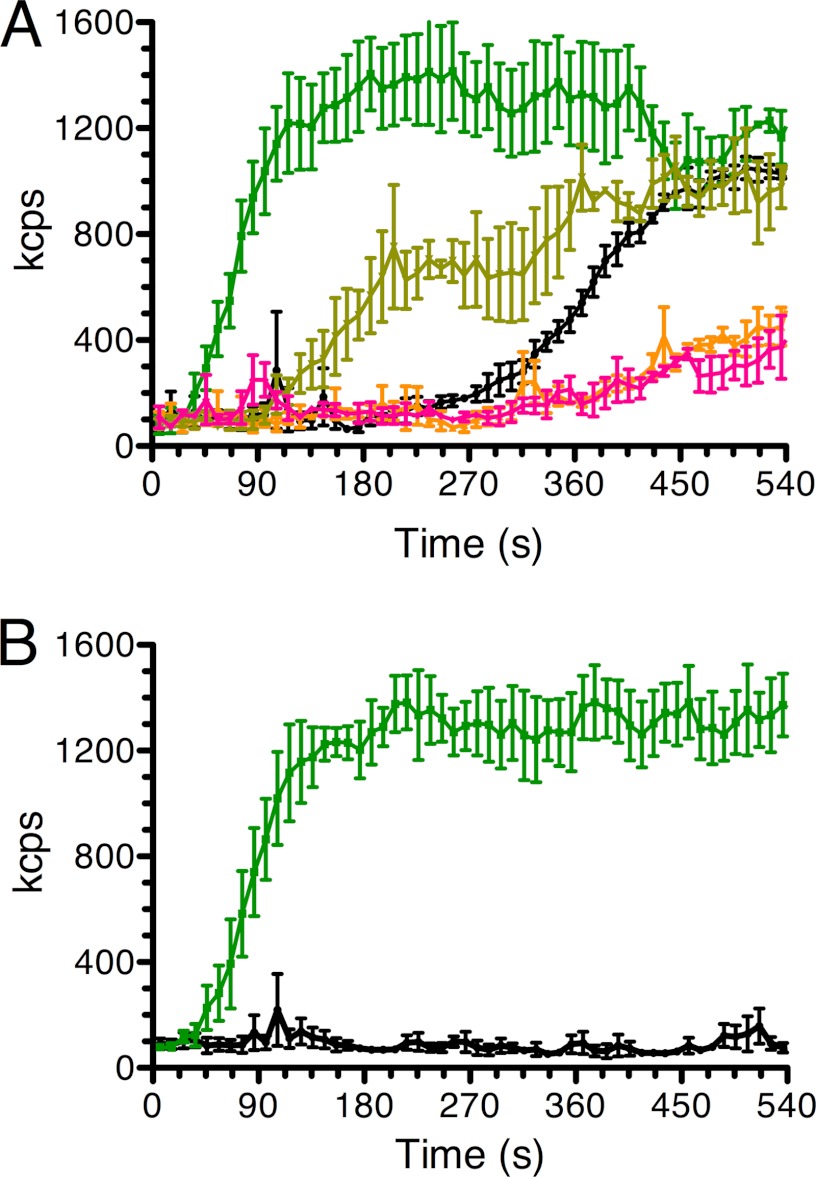
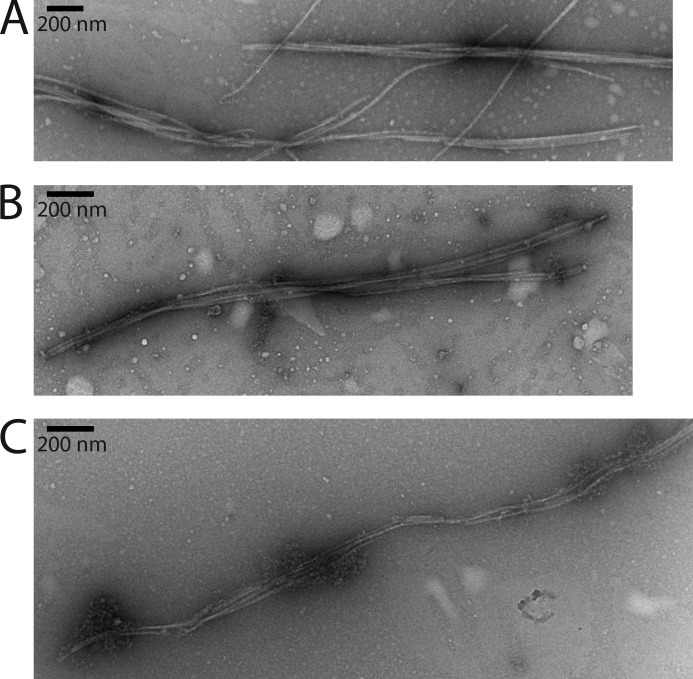
The actin homolog MreB is required in rod-shaped bacteria for maintenance of cell shape and is intimately connected to the holoenzyme that synthesizes the peptidoglycan layer. The protein has been reported variously to exist in helical loops under the cell surface, to rotate, and to move in patches in both directions around the cell surface. Studies of the Escherichia coli protein in vitro have been hampered by its tendency to aggregate. Here we report the purification and characterization of native E. coli MreB. The protein requires ATP hydrolysis for polymerization, forms bundles with a left-hand twist that can be as long as 4 μm, forms sheets in the presence of calcium, and has a critical concentration for polymerization of 1.5 μM.
Figures
References
-
- Jones L. J., Carballido-López R., Errington J. (2001) Control of cell shape in bacteria: helical, actin-like filaments in Bacillus subtilis. Cell 104, 913–922 - PubMed
-
- van den Ent F., Amos L. A., Löwe J. (2001) Prokaryotic origin of the actin cytoskeleton. Nature 413, 39–44 - PubMed
-
- Møller-Jensen J., Borch J., Dam M., Jensen R. B., Roepstorff P., Gerdes K. (2003) Bacterial mitosis: ParM of plasmid R1 moves plasmid DNA by an actin-like insertional polymerization mechanism. Mol. Cell 12, 1477–1487 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
