

The anti-proliferative activity of BTG/TOB proteins is mediated via the Caf1a (CNOT7) and Caf1b (CNOT8) deadenylase subunits of the Ccr4-not complex
- PMID: 23236473
- PMCID: PMC3517456
- DOI: 10.1371/journal.pone.0051331
The anti-proliferative activity of BTG/TOB proteins is mediated via the Caf1a (CNOT7) and Caf1b (CNOT8) deadenylase subunits of the Ccr4-not complex
Abstract
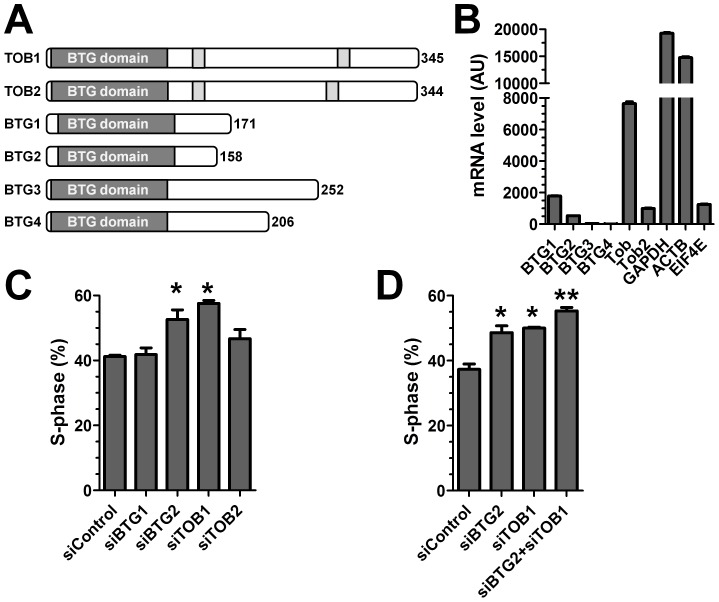
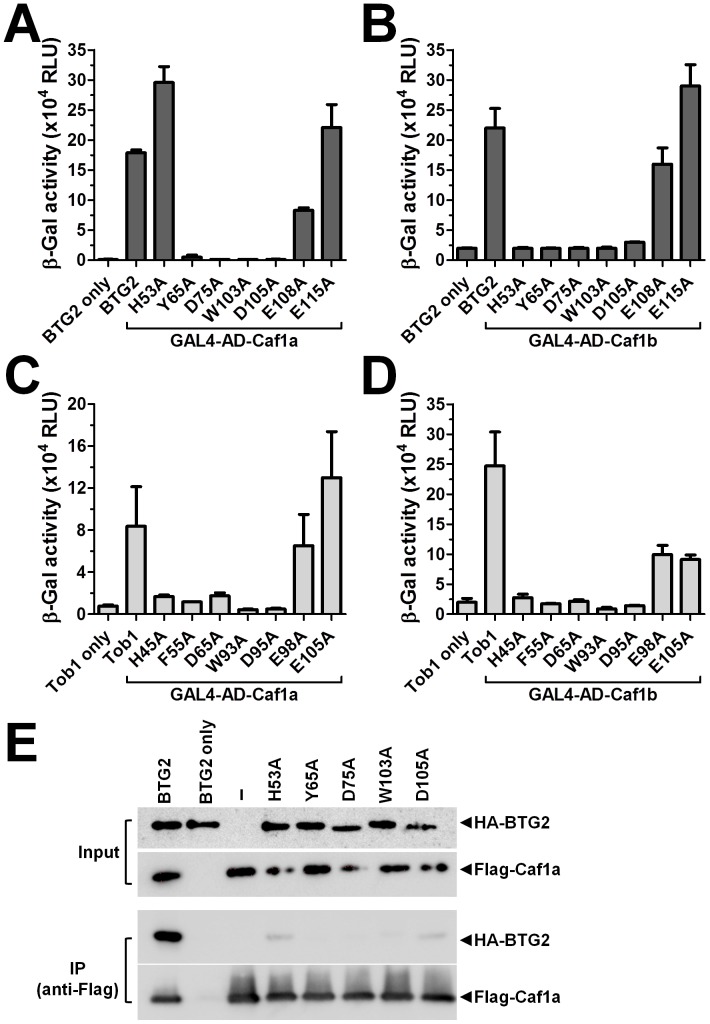
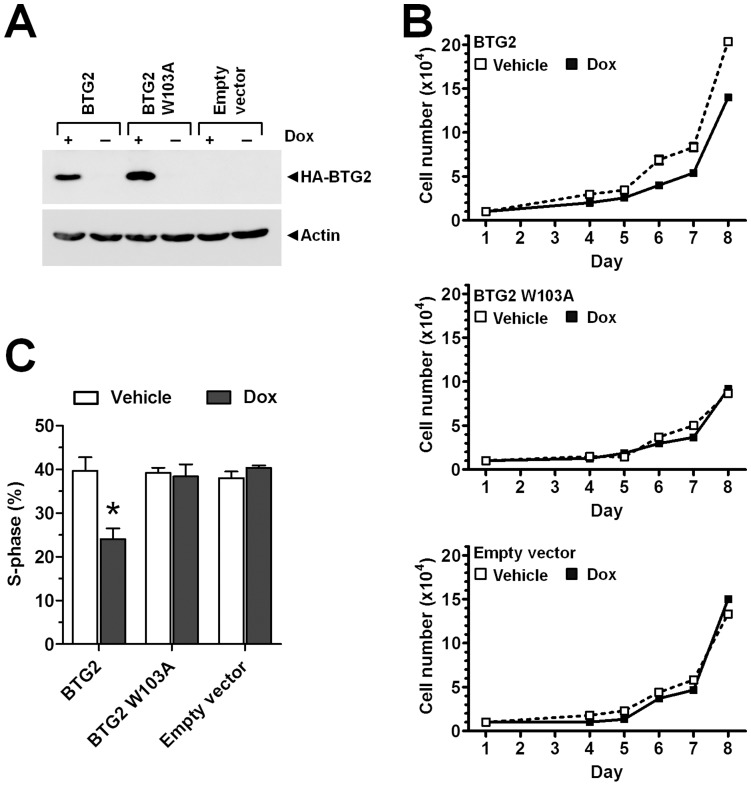
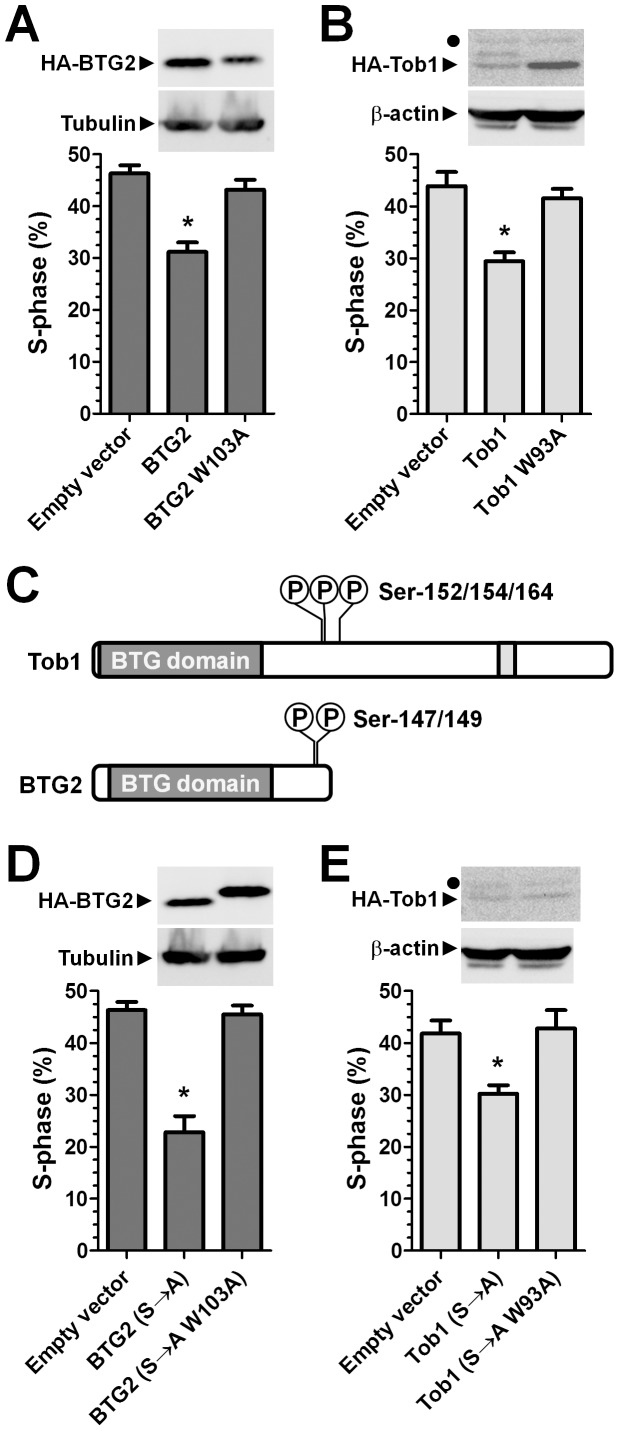
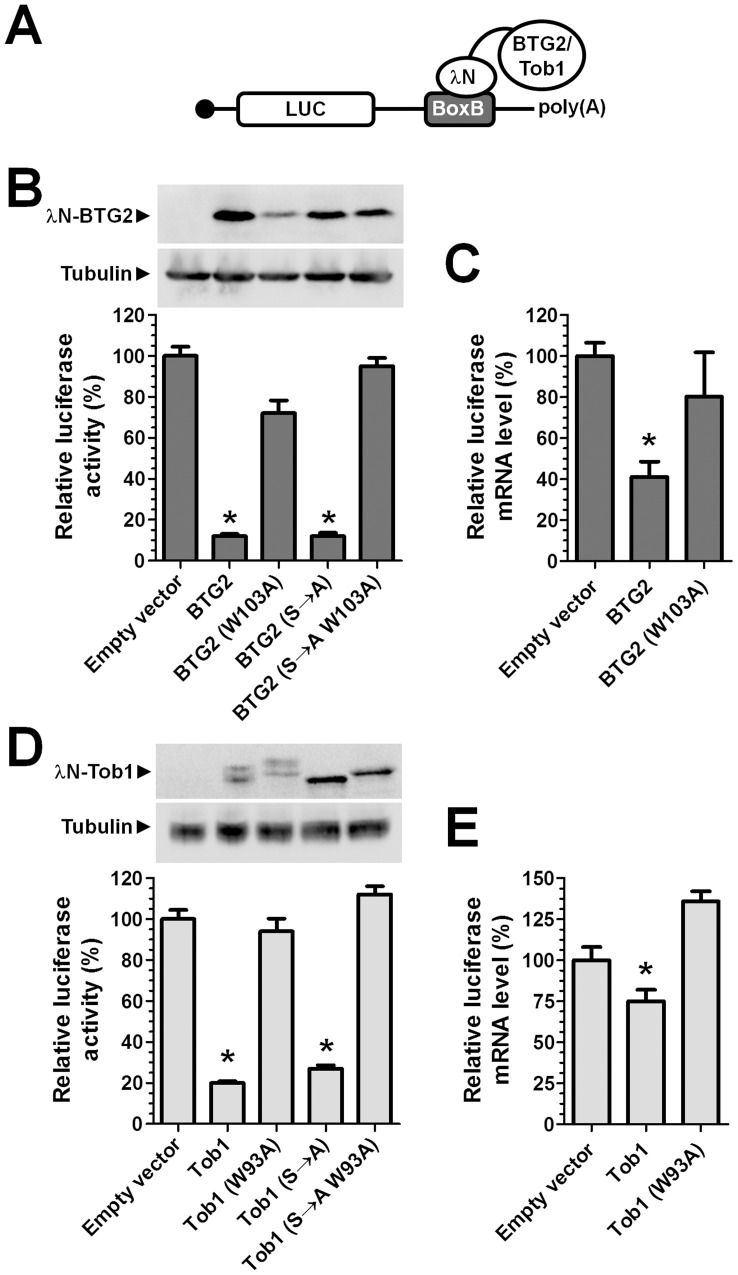
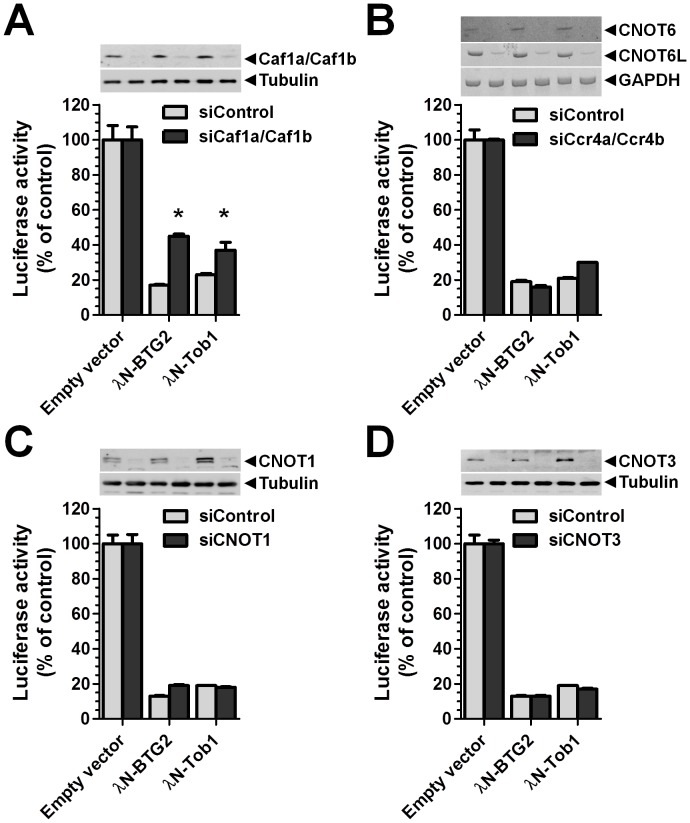
The human BTG/TOB protein family comprises six members (BTG1, BTG2/PC3/Tis21, BTG3/Ana, BTG4/PC3B, TOB1/Tob, and TOB2) that are characterised by a conserved BTG domain. This domain mediates interactions with the highly similar Caf1a (CNOT7) and Caf1b (CNOT8) catalytic subunits of the Ccr4-Not deadenylase complex. BTG/TOB proteins have anti-proliferative activity: knockdown of BTG/TOB can result in increased cell proliferation, whereas over-expression of BTG/TOB leads to inhibition of cell cycle progression. It was unclear whether the interaction between BTG/TOB proteins and the Caf1a/Caf1b deadenylases is necessary for the anti-proliferative activity of BTG/TOB. To address this question, we further characterised surface-exposed amino acid residues of BTG2 and TOB1 that mediate the interaction with the Caf1a and Caf1b deadenylase enzymes. We then analysed the role of BTG2 and TOB1 in the regulation of cell proliferation, translation and mRNA abundance using a mutant that is no longer able to interact with the Caf1a/Caf1b deadenylases. We conclude that the anti-proliferative activity of BTG/TOB proteins is mediated through interactions with the Caf1a and Caf1b deadenylase enzymes. Furthermore, we show that the activity of BTG/TOB proteins in the regulation of mRNA abundance and translation is dependent on Caf1a/Caf1b, and does not appear to require other Ccr4-Not components, including the Ccr4a (CNOT6)/Ccr4b (CNOT6L) deadenylases, or the non-catalytic subunits CNOT1 or CNOT3.
Conflict of interest statement
Figures
Similar articles
-
The mammalian anti-proliferative BTG/Tob protein family.J Cell Physiol. 2010 Jan;222(1):66-72. doi: 10.1002/jcp.21919. J Cell Physiol. 2010. PMID: 19746446 Review.
-
The Ccr4a (CNOT6) and Ccr4b (CNOT6L) deadenylase subunits of the human Ccr4-Not complex contribute to the prevention of cell death and senescence.Mol Biol Cell. 2011 Mar 15;22(6):748-58. doi: 10.1091/mbc.E10-11-0898. Epub 2011 Jan 13. Mol Biol Cell. 2011. PMID: 21233283 Free PMC article.
-
The Ccr4-NOT deadenylase subunits CNOT7 and CNOT8 have overlapping roles and modulate cell proliferation.Mol Biol Cell. 2009 Sep;20(17):3840-50. doi: 10.1091/mbc.e09-02-0146. Epub 2009 Jul 15. Mol Biol Cell. 2009. PMID: 19605561 Free PMC article.
-
BTG/Tob family members Tob1 and Tob2 inhibit proliferation of mouse embryonic stem cells via Id3 mRNA degradation.Biochem Biophys Res Commun. 2015 Jul 3;462(3):208-14. doi: 10.1016/j.bbrc.2015.04.117. Epub 2015 May 5. Biochem Biophys Res Commun. 2015. PMID: 25951976
-
Deadenylation of cytoplasmic mRNA by the mammalian Ccr4-Not complex.Biochem Soc Trans. 2012 Aug;40(4):896-901. doi: 10.1042/BST20120074. Biochem Soc Trans. 2012. PMID: 22817755 Review.
Cited by
-
Role of the Citrus sinensis RNA deadenylase CsCAF1 in citrus canker resistance.Mol Plant Pathol. 2019 Aug;20(8):1105-1118. doi: 10.1111/mpp.12815. Epub 2019 May 21. Mol Plant Pathol. 2019. PMID: 31115151 Free PMC article.
-
CircRNAs and RNA-Binding Proteins Involved in the Pathogenesis of Cancers or Central Nervous System Disorders.Noncoding RNA. 2023 Mar 31;9(2):23. doi: 10.3390/ncrna9020023. Noncoding RNA. 2023. PMID: 37104005 Free PMC article. Review.
-
The Ccr4-Not complex is a key regulator of eukaryotic gene expression.Wiley Interdiscip Rev RNA. 2016 Jul;7(4):438-54. doi: 10.1002/wrna.1332. Epub 2016 Jan 29. Wiley Interdiscip Rev RNA. 2016. PMID: 26821858 Free PMC article. Review.
-
Identification of biomarkers for endometriosis based on summary-data-based Mendelian randomization and machine learning.Medicine (Baltimore). 2025 Apr 4;104(14):e41804. doi: 10.1097/MD.0000000000041804. Medicine (Baltimore). 2025. PMID: 40193647 Free PMC article.
-
Tob2 phosphorylation regulates global mRNA turnover to reshape transcriptome and impact cell proliferation.RNA. 2020 Sep;26(9):1143-1159. doi: 10.1261/rna.073528.119. Epub 2020 May 13. RNA. 2020. PMID: 32404348 Free PMC article.
References
-
- Winkler GS (2010) The mammalian anti-proliferative BTG/Tob protein family. J Cell Physiol 222: 66–72. - PubMed
-
- Okochi K, Suzuki T, Inoue J, Matsuda S, Yamamoto T (2005) Interaction of anti-proliferative protein Tob with poly(A)-binding protein and inducible poly(A)-binding protein: implication of Tob in translational control. Genes Cells 10: 151–163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
