

Neto2 interacts with the scaffolding protein GRIP and regulates synaptic abundance of kainate receptors
- PMID: 23236500
- PMCID: PMC3516556
- DOI: 10.1371/journal.pone.0051433
Neto2 interacts with the scaffolding protein GRIP and regulates synaptic abundance of kainate receptors
Abstract
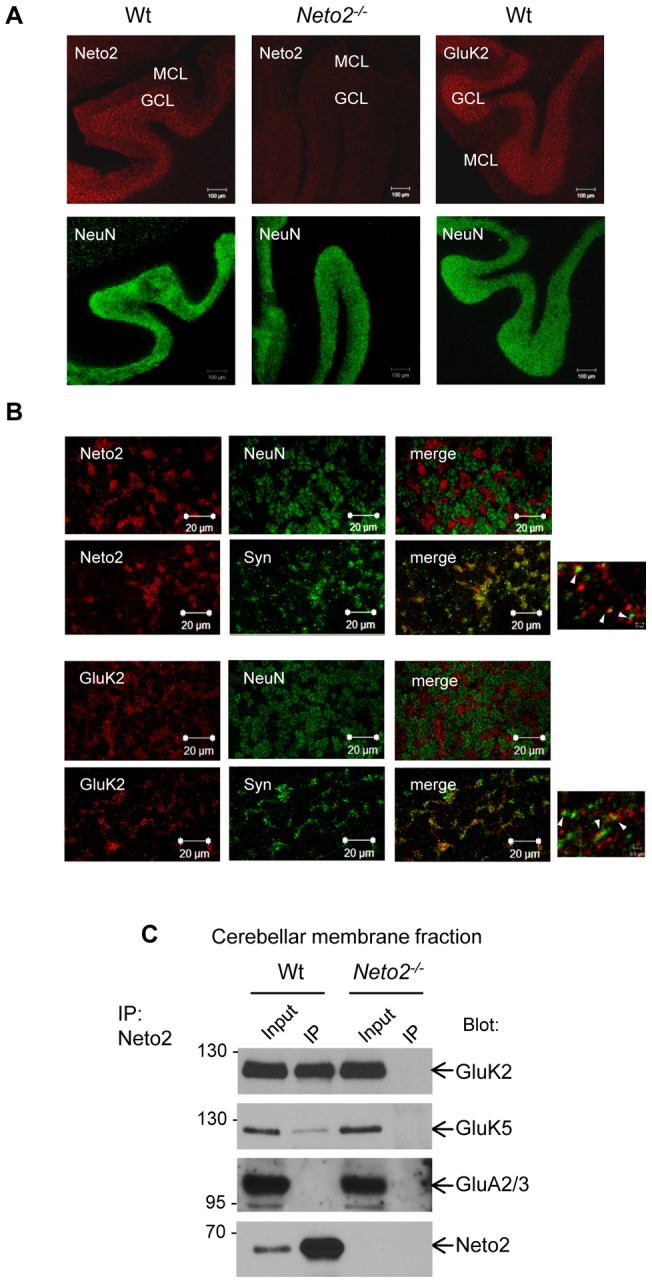
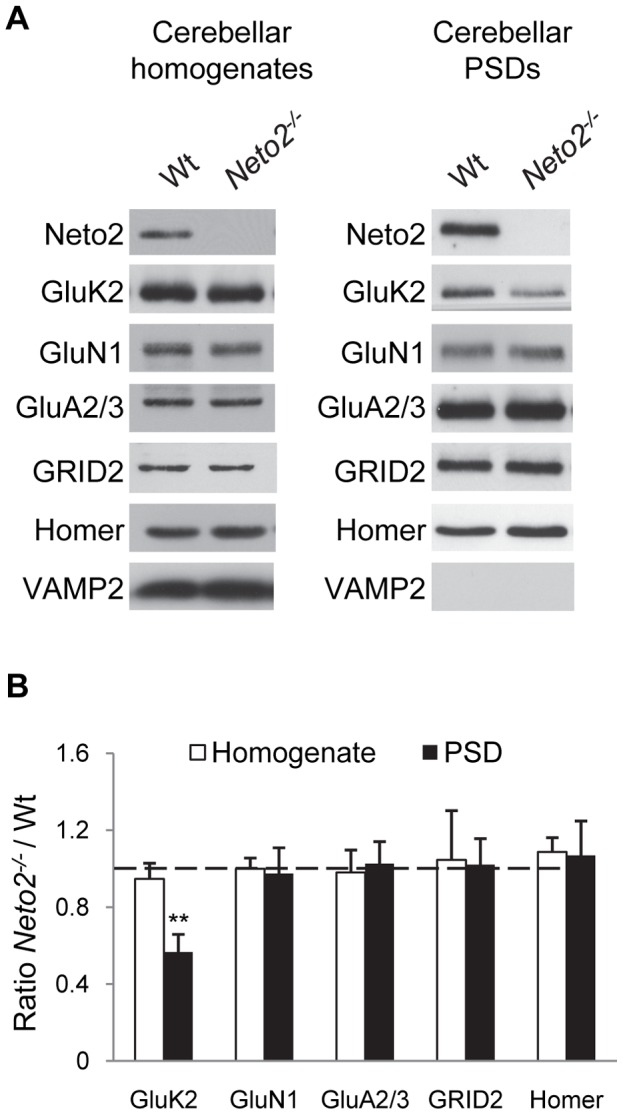
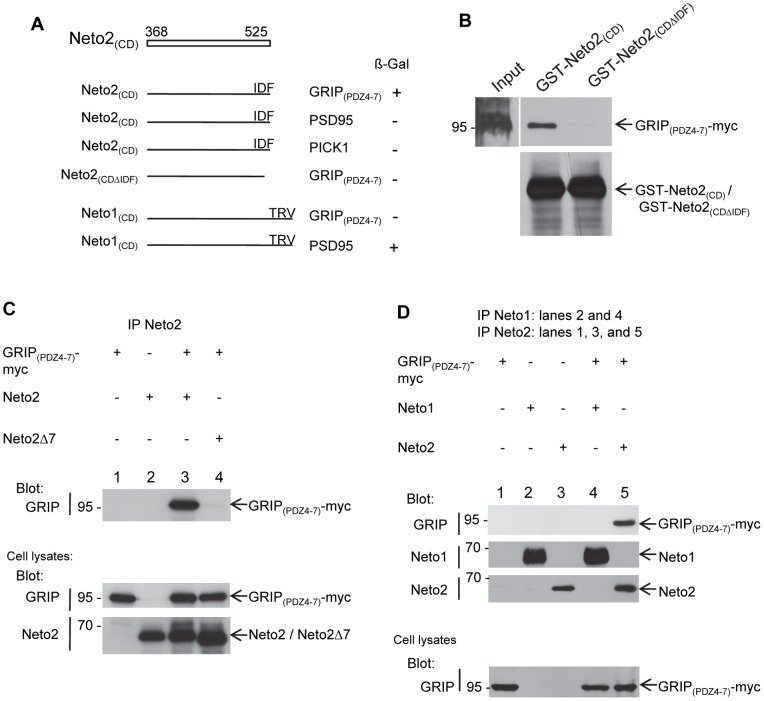
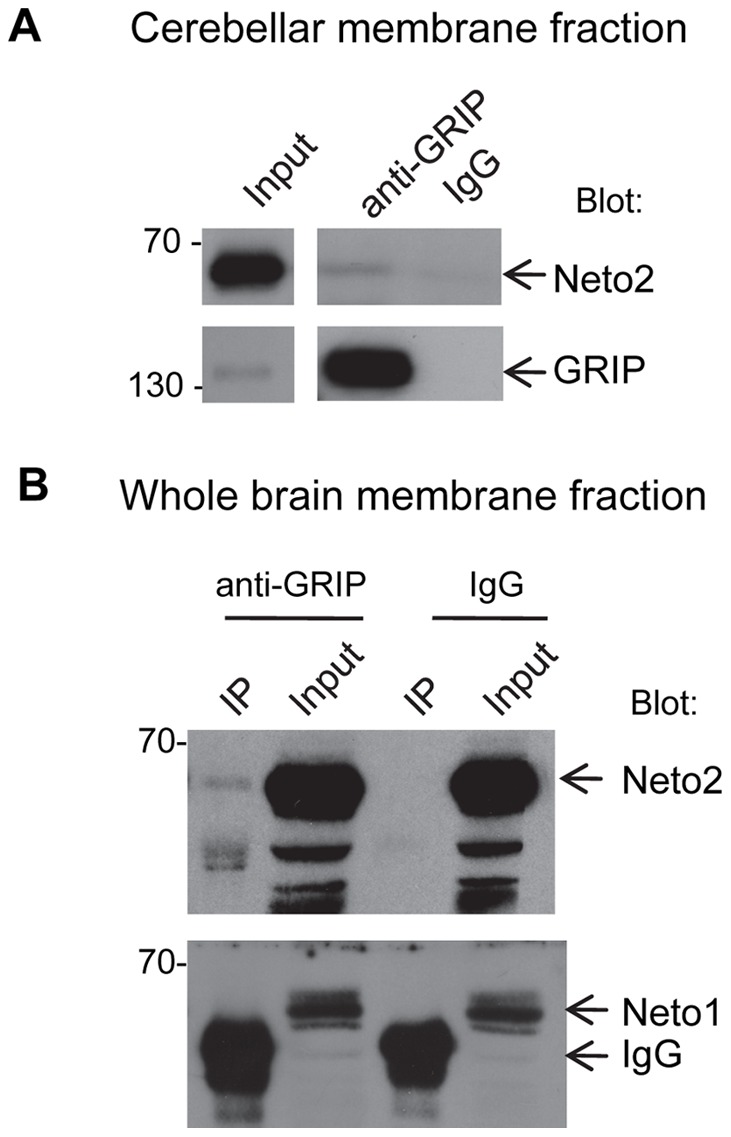
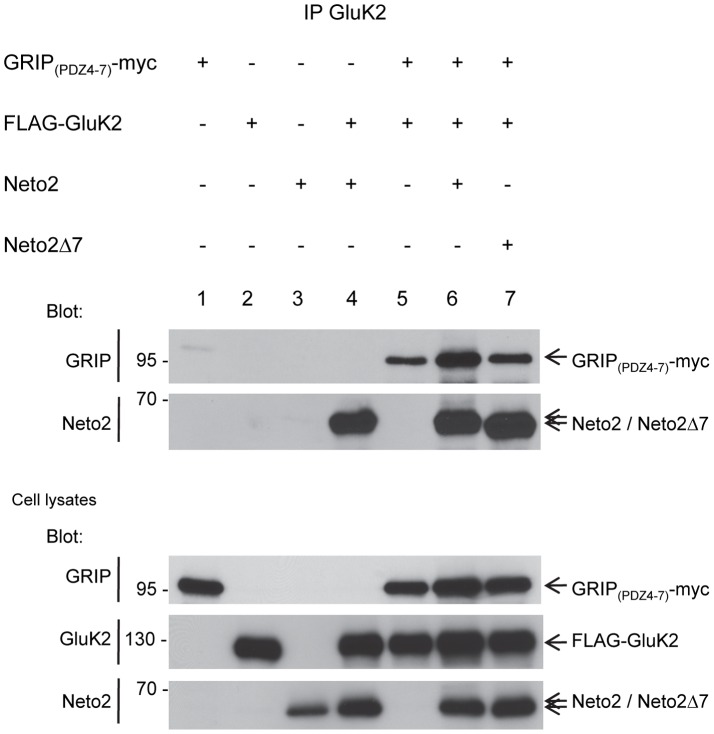
Kainate receptors (KARs) are a class of ionotropic glutamate receptors that are expressed throughout the central nervous system. The function and subcellular localization of KARs are tightly regulated by accessory proteins. We have previously identified the single-pass transmembrane proteins, Neto1 and Neto2, to be associated with native KARs. In the hippocampus, Neto1, but not Neto2, controls the abundance and modulates the kinetics of postsynaptic KARs. Here we evaluated whether Neto2 regulates synaptic KAR levels in the cerebellum where Neto1 expression is limited to the deep cerebellar nuclei. In the cerebellum, where Neto2 is present abundantly, we found a ~40% decrease in GluK2-KARs at the postsynaptic density (PSD) of Neto2-null mice. No change, however, was observed in total level of GluK2-KARs, thereby suggesting a critical role of Neto2 on the synaptic localization of cerebellar KARs. The presence of a putative class II PDZ binding motif on Neto2 led us to also investigate whether it interacts with PDZ domain-containing proteins previously implicated in regulating synaptic abundance of KARs. We identified a PDZ-dependent interaction between Neto2 and the scaffolding protein GRIP. Furthermore, coexpression of Neto2 significantly increased the amount of GRIP associated with GluK2, suggesting that Neto2 may promote and/or stabilize GluK2:GRIP interactions. Our results demonstrate that Neto2, like Neto1, is an important auxiliary protein for modulating the synaptic levels of KARs. Moreover, we propose that the interactions of Neto1/2 with various scaffolding proteins is a critical mechanism by which KARs are stabilized at diverse synapses.
Conflict of interest statement
Figures
References
-
- Pinheiro PS, Mulle C (2008) Presynaptic glutamate receptors: physiological functions and mechanisms of action. Nat Rev Neurosci 9: 423–436. - PubMed
-
- Castillo PE, Malenka RC, Nicoll RA (1997) Kainate receptors mediate a slow postsynaptic current in hippocampal CA3 neurons. Nature 388: 182–186. - PubMed
-
- Vignes M, Collingridge GL (1997) The synaptic activation of kainate receptors. Nature 388: 179–182. - PubMed
-
- Cossart R, Esclapez M, Hirsch JC, Bernard C, Ben-Ari Y (1998) GluR5 kainate receptor activation in interneurons increases tonic inhibition of pyramidal cells. Nat Neurosci 1: 470–478. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
