

Bacillus anthracis acetyltransferases PatA1 and PatA2 modify the secondary cell wall polysaccharide and affect the assembly of S-layer proteins
- PMID: 23243307
- PMCID: PMC3571321
- DOI: 10.1128/JB.01274-12
Bacillus anthracis acetyltransferases PatA1 and PatA2 modify the secondary cell wall polysaccharide and affect the assembly of S-layer proteins
Abstract
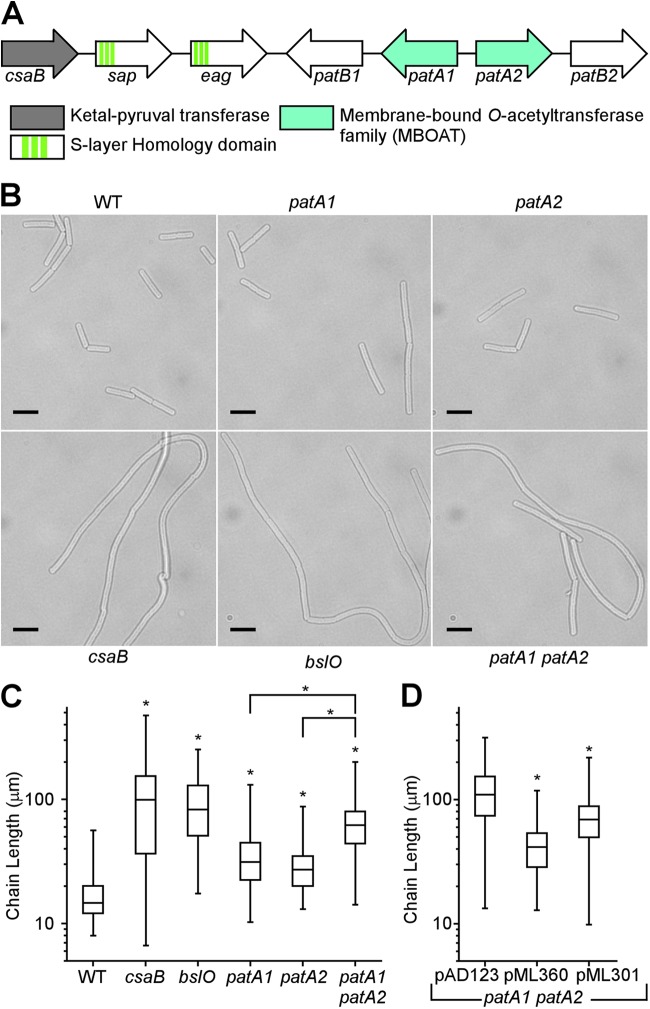
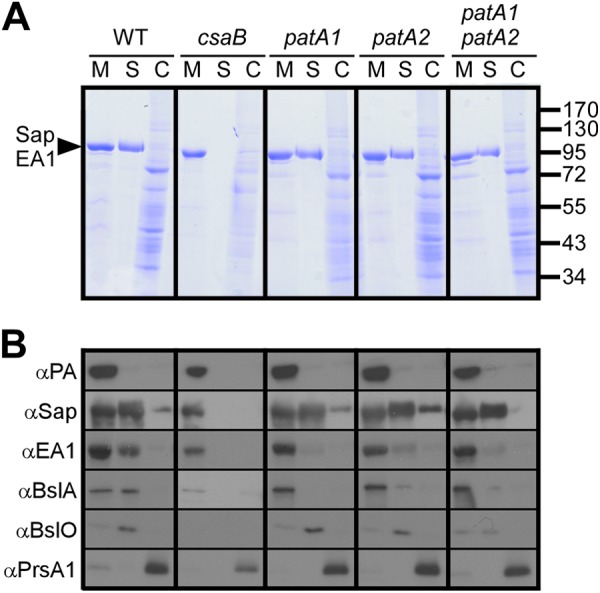
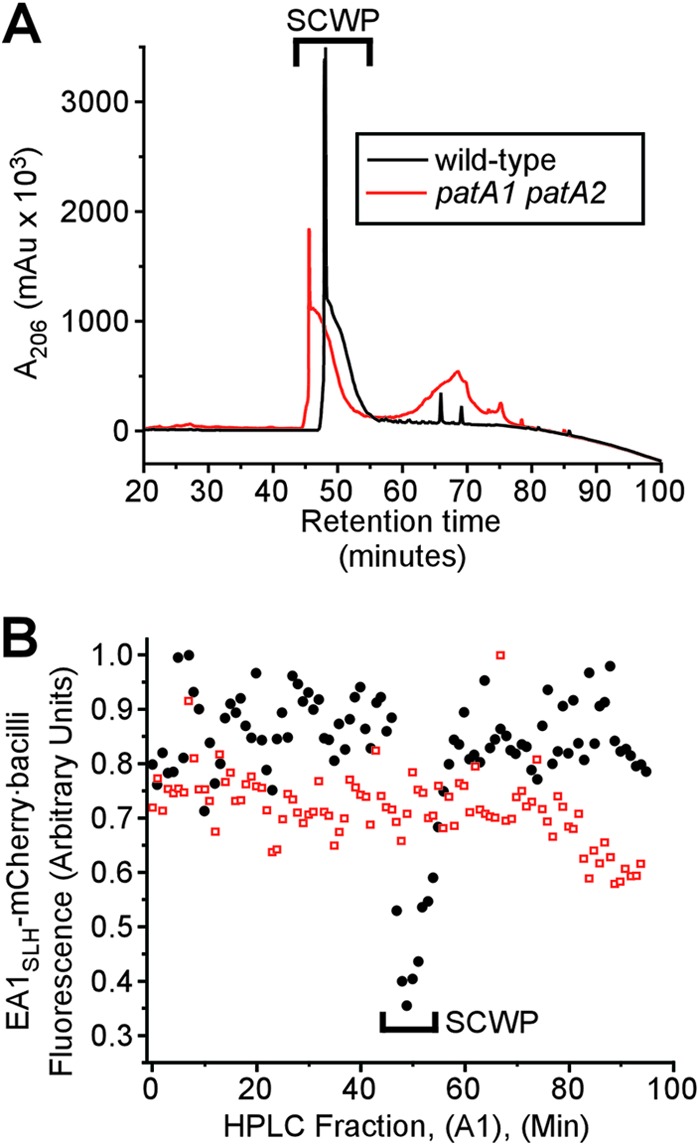
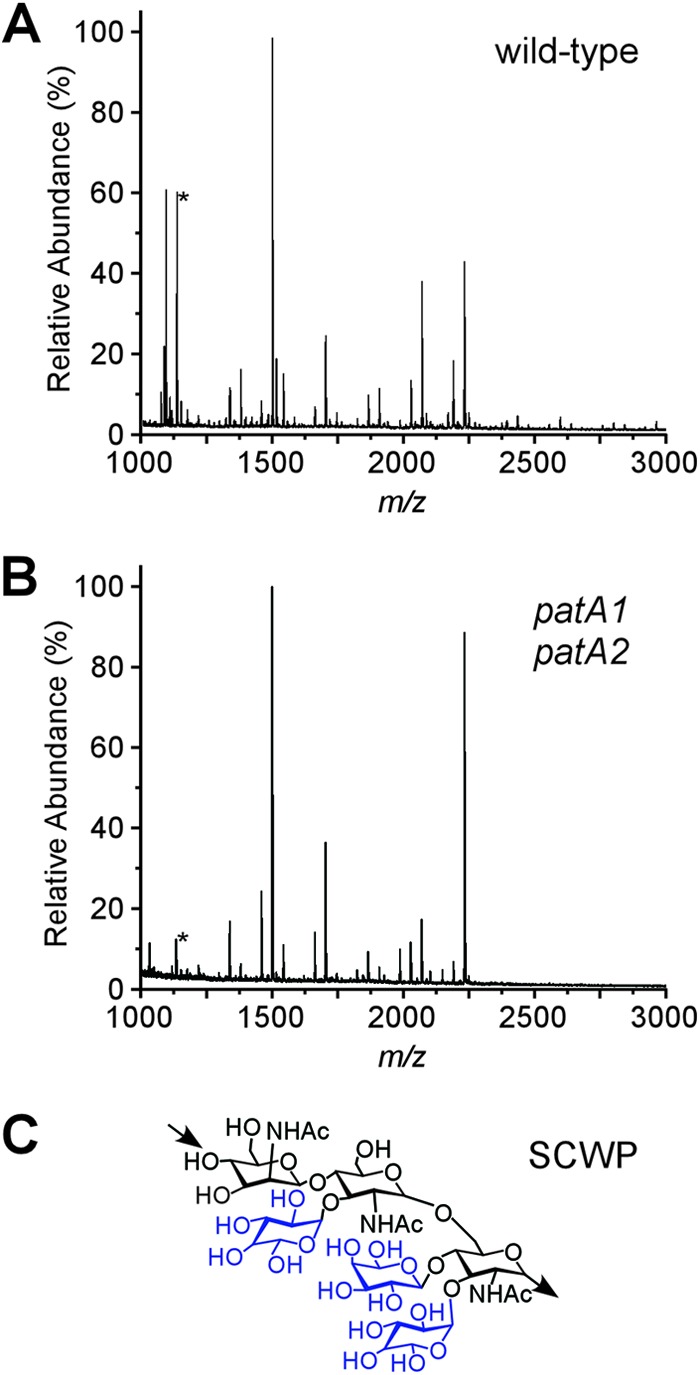
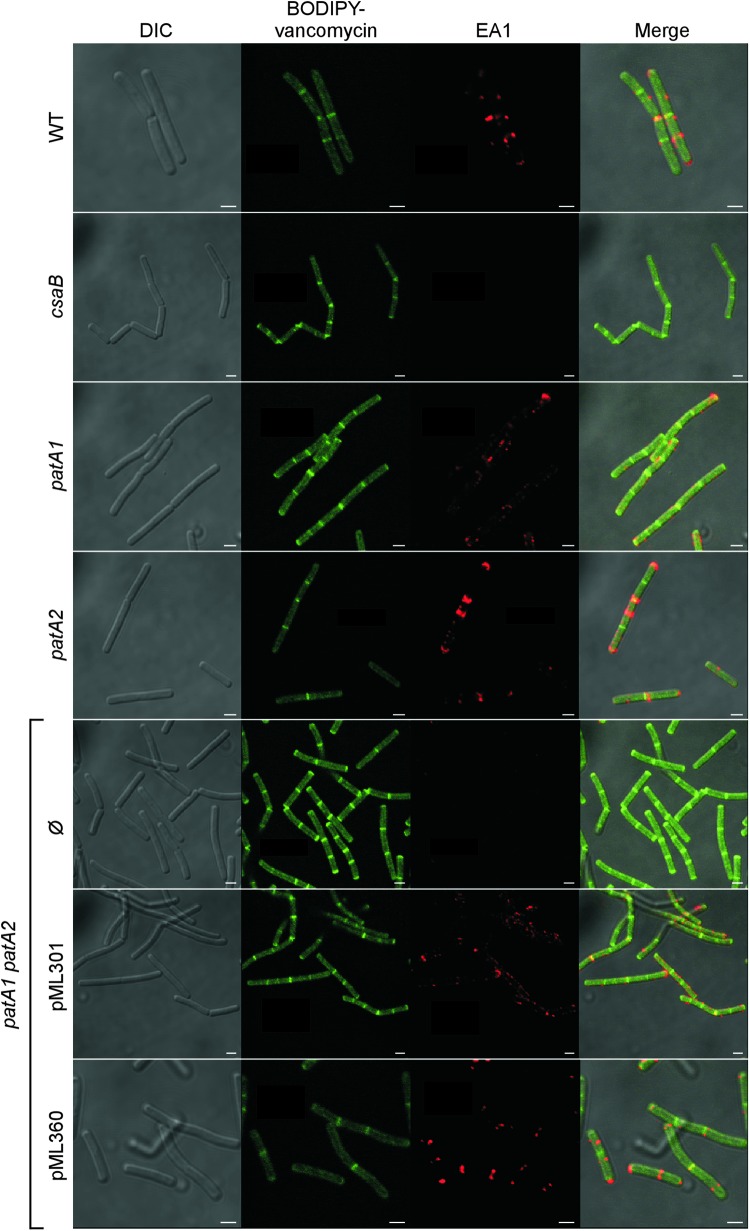
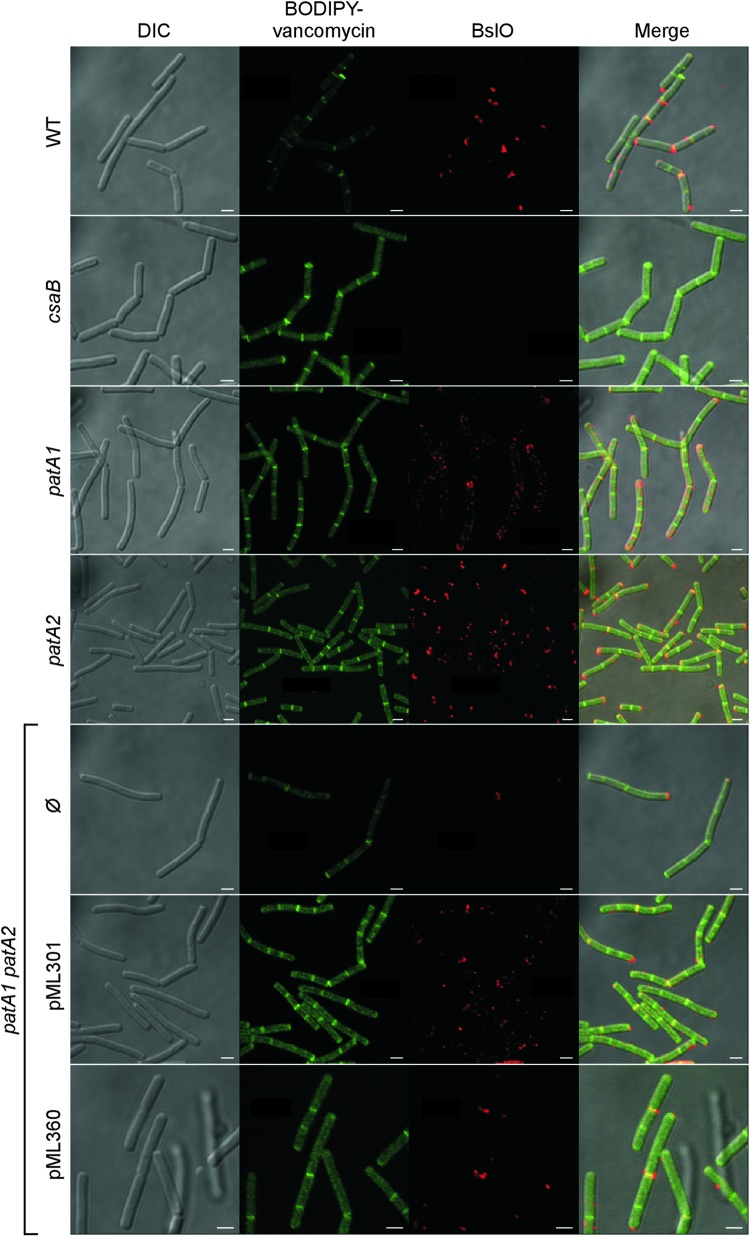
The envelope of Bacillus anthracis encompasses a proteinaceous S-layer with two S-layer proteins (Sap and EA1). Protein assembly in the envelope of B. anthracis requires S-layer homology domains (SLH) within S-layer proteins and S-layer-associated proteins (BSLs), which associate with the secondary cell wall polysaccharide (SCWP), an acetylated carbohydrate that is tethered to peptidoglycan. Here, we investigated the contributions of two putative acetyltransferases, PatA1 and PatA2, on SCWP acetylation and S-layer assembly. We show that mutations in patA1 and patA2 affect the chain lengths of B. anthracis vegetative forms and perturb the deposition of the BslO murein hydrolase at cell division septa. The patA1 and patA2 mutants are defective for the assembly of EA1 in the envelope but retain the ability of S-layer formation with Sap. SCWP isolated from the patA1 patA2 mutant lacked acetyl moieties identified in wild-type polysaccharide and failed to associate with the SLH domains of EA1. A model is discussed whereby patA1- and patA2-mediated acetylation of SCWP enables the deposition of EA1 as well as BslO near the septal region of the B. anthracis envelope.
Figures
References
-
- Koch R. 1876. Die Ätiologie der Milzbrand-Krankheit, begründet auf die Entwicklungsgeschichte des Bacillus anthracis. Beitr. Biol. Pflanz. 2: 277– 310
-
- Marraffini LA, Schneewind O. 2006. Targeting proteins to the cell wall of sporulating Bacillus anthracis. Mol. Microbiol. 62: 1402– 1417 - PubMed
-
- Candela T, Fouet A. 2006. Poly-gamma-glutamate in bacteria. Mol. Microbiol. 60: 1091– 1098 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
