

Ribonucleotide incorporation, proofreading and bypass by human DNA polymerase δ
- PMID: 23245697
- PMCID: PMC3552135
- DOI: 10.1016/j.dnarep.2012.11.006
Ribonucleotide incorporation, proofreading and bypass by human DNA polymerase δ
Abstract
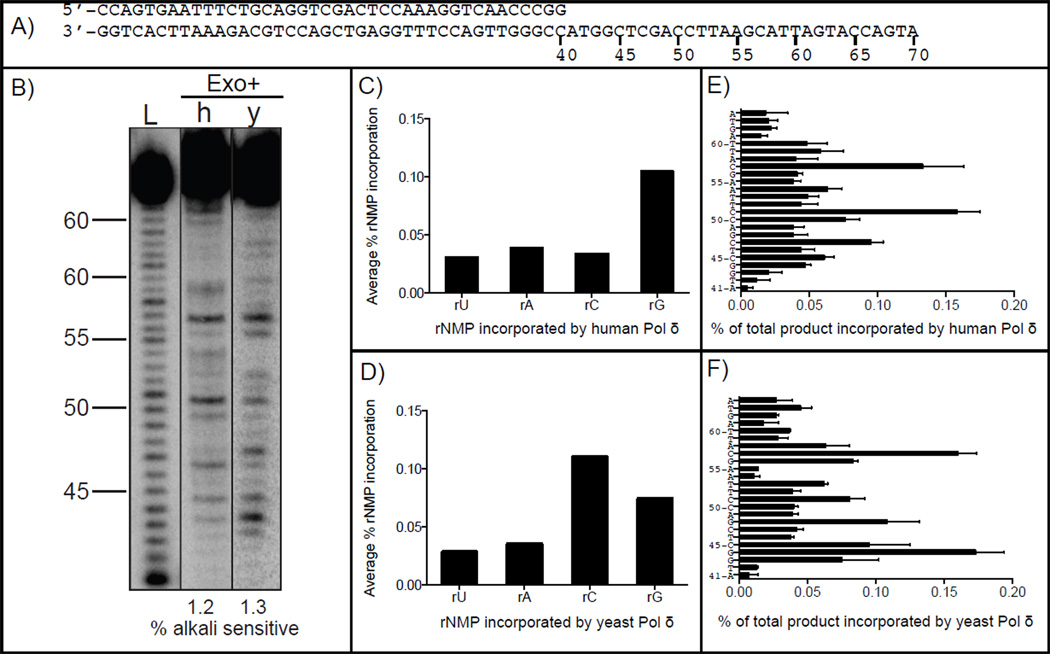
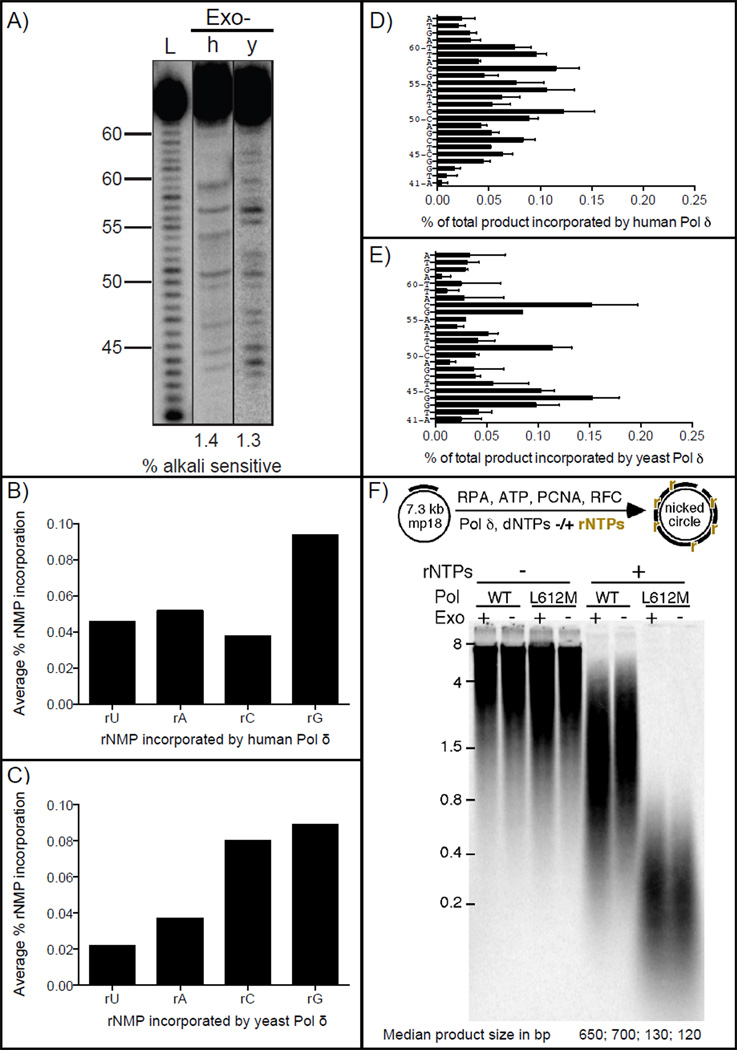
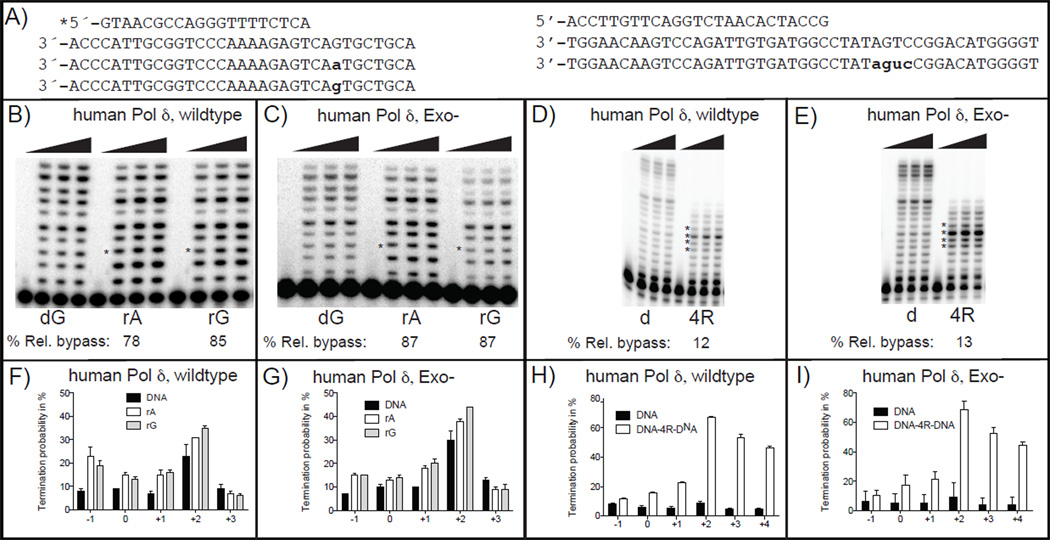
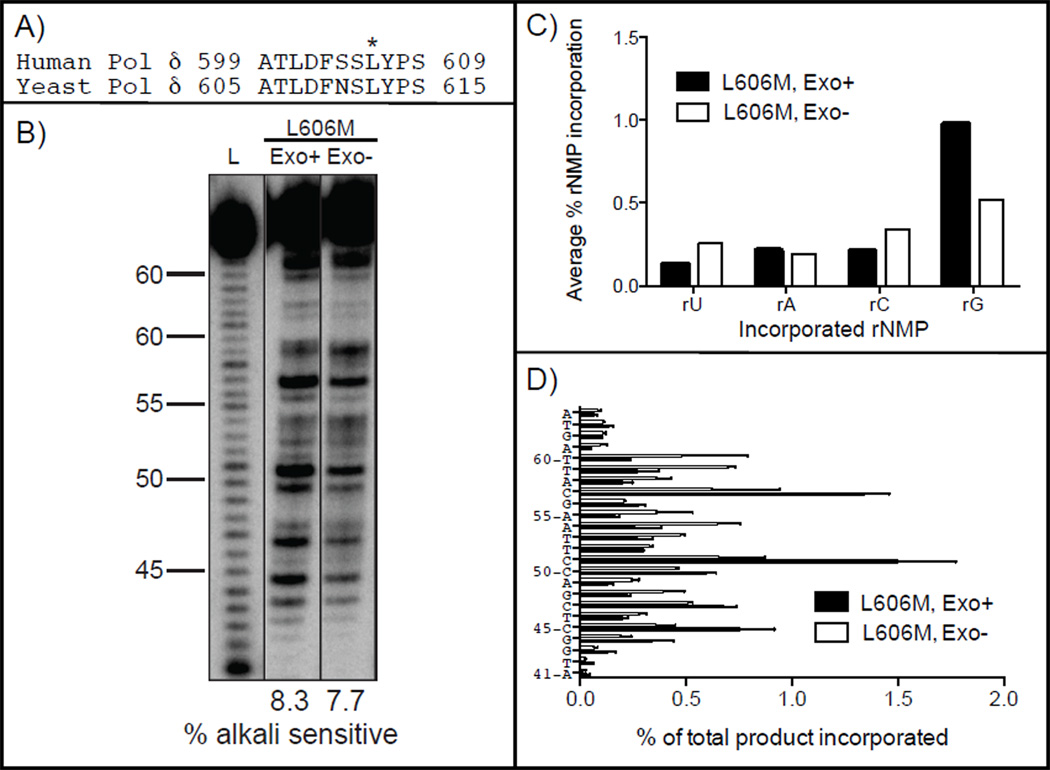
In both budding and fission yeast, a large number of ribonucleotides are incorporated into DNA during replication by the major replicative polymerases (Pols α, δ and ɛ). They are subsequently removed by RNase H2-dependent repair, which if defective leads to replication stress and genome instability. To extend these studies to humans, where an RNase H2 defect results in an autoimmune disease, here we compare the ability of human and yeast Pol δ to incorporate, proofread, and bypass ribonucleotides during DNA synthesis. In reactions containing nucleotide concentrations estimated to be present in mammalian cells, human Pol δ stably incorporates one rNTP for approximately 2000 dNTPs, a ratio similar to that for yeast Pol δ. This result predicts that human Pol δ may introduce more than a million ribonucleotides into the nuclear genome per replication cycle, an amount recently reported to be present in the genome of RNase H2-defective mouse cells. Consistent with such abundant stable incorporation, we show that the 3'-exonuclease activity of yeast and human Pol δ largely fails to edit ribonucleotides during polymerization. We also show that, like yeast Pol δ, human Pol δ pauses as it bypasses ribonucleotides in DNA templates, with four consecutive ribonucleotides in a DNA template being more problematic than single ribonucleotides. In conjunction with recent studies in yeast and mice, this ribonucleotide incorporation may be relevant to impaired development and disease when RNase H2 is defective in mammals. As one tool to investigate ribonucleotide incorporation by Pol δ in human cells, we show that human Pol δ containing a Leu606Met substitution in the polymerase active site incorporates 7-fold more ribonucleotides into DNA than does wild type Pol δ.
Published by Elsevier B.V.
Figures
References
-
- Kornberg A, Baker TA. DNA Replication. Herndon, Virginia: University Science Books; 2005.
-
- Traut TW. Physiological concentrations of purines and pyrimidines. Mol. Cell. Biochem. 1994;140:1–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
