

Mutations in the β-tubulin gene TUBB5 cause microcephaly with structural brain abnormalities
- PMID: 23246003
- PMCID: PMC3595605
- DOI: 10.1016/j.celrep.2012.11.017
Mutations in the β-tubulin gene TUBB5 cause microcephaly with structural brain abnormalities
Abstract
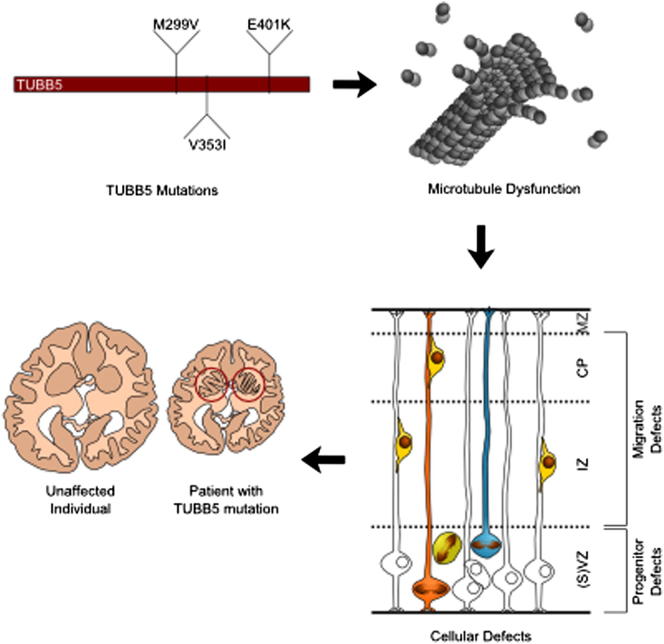
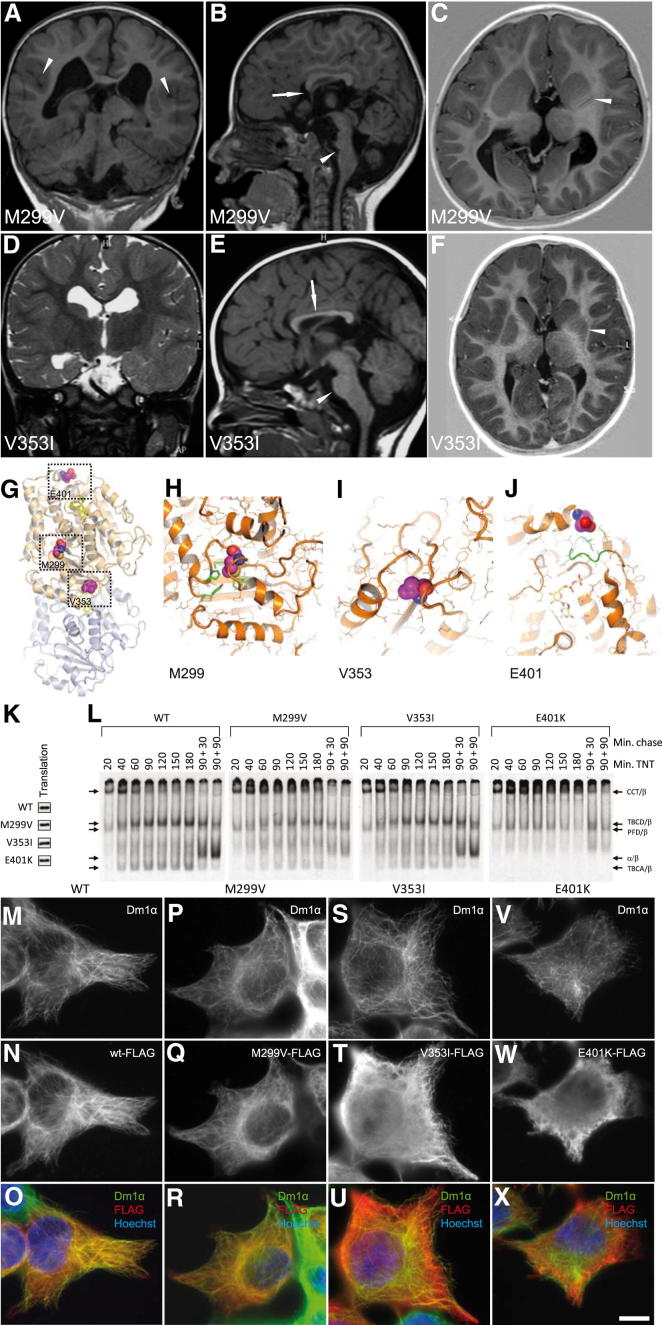
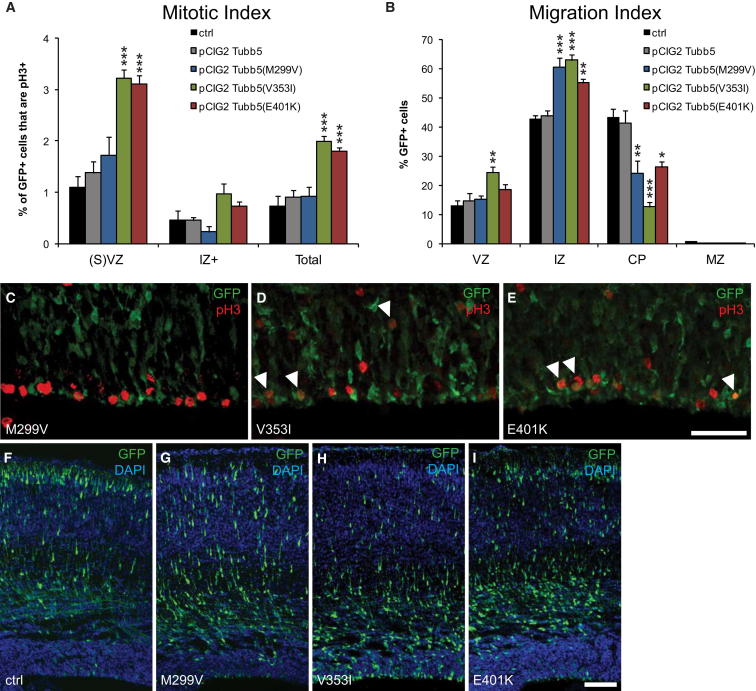
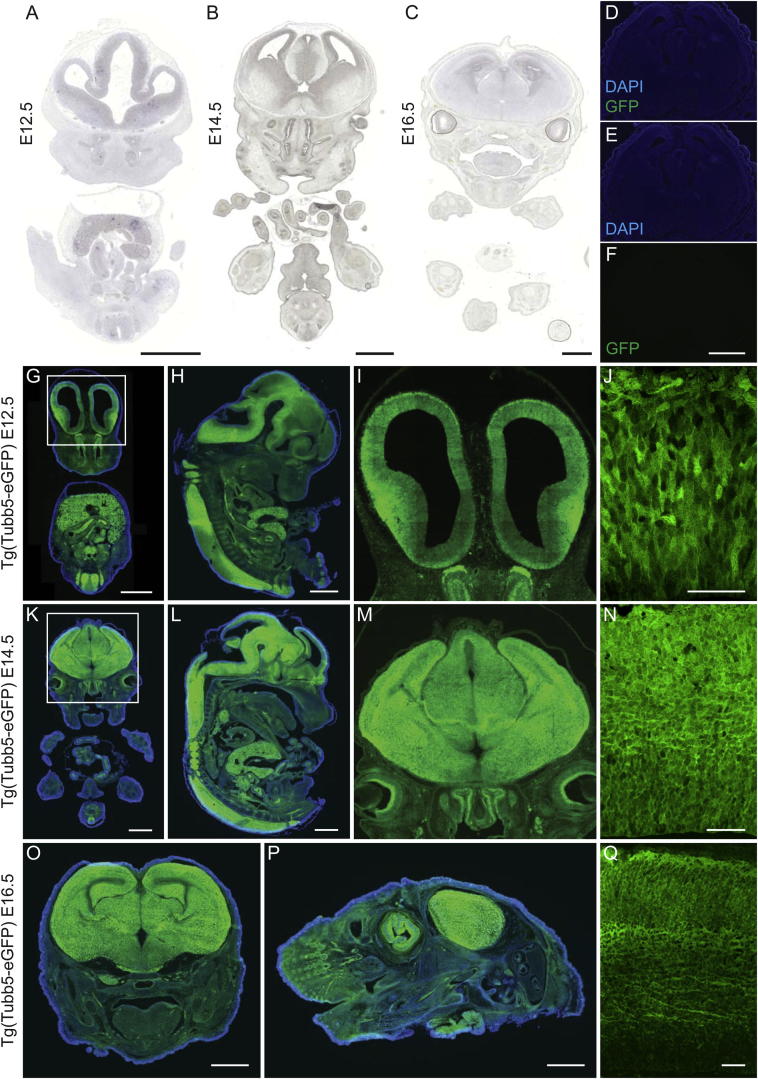
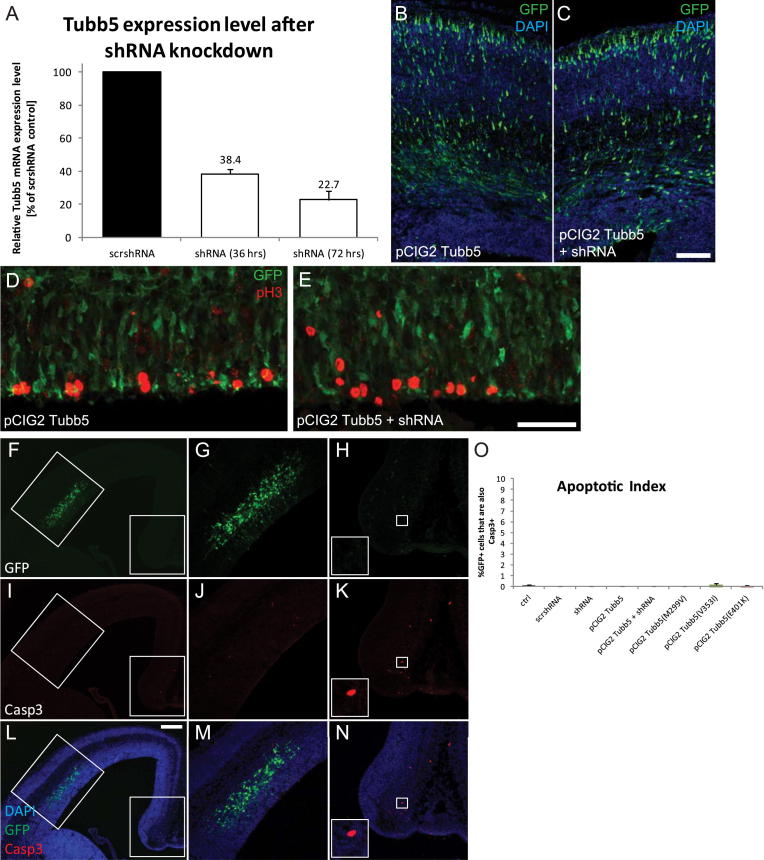
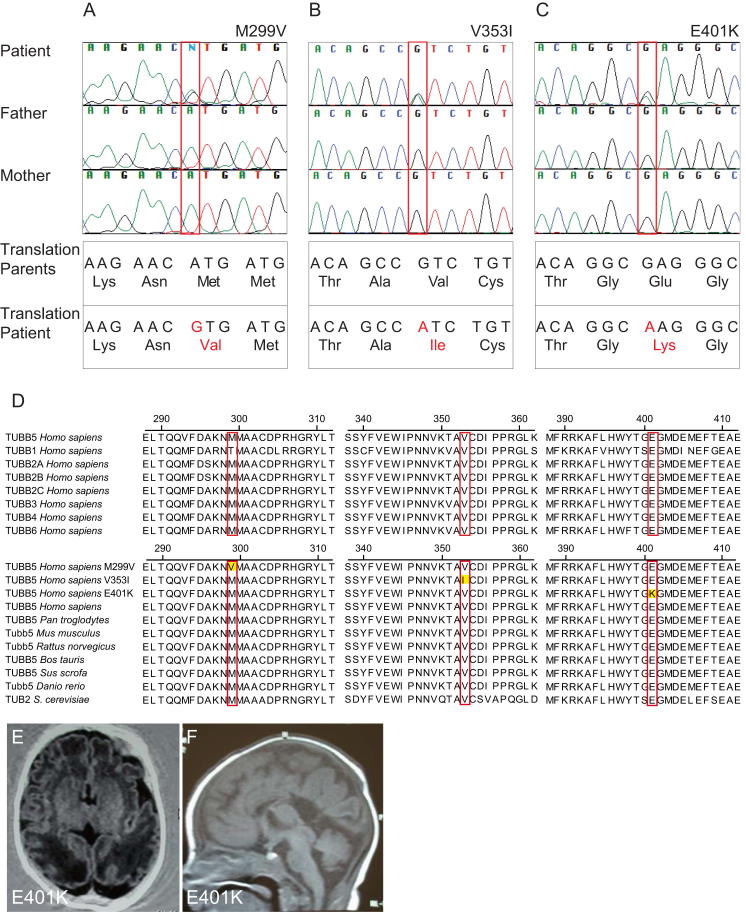
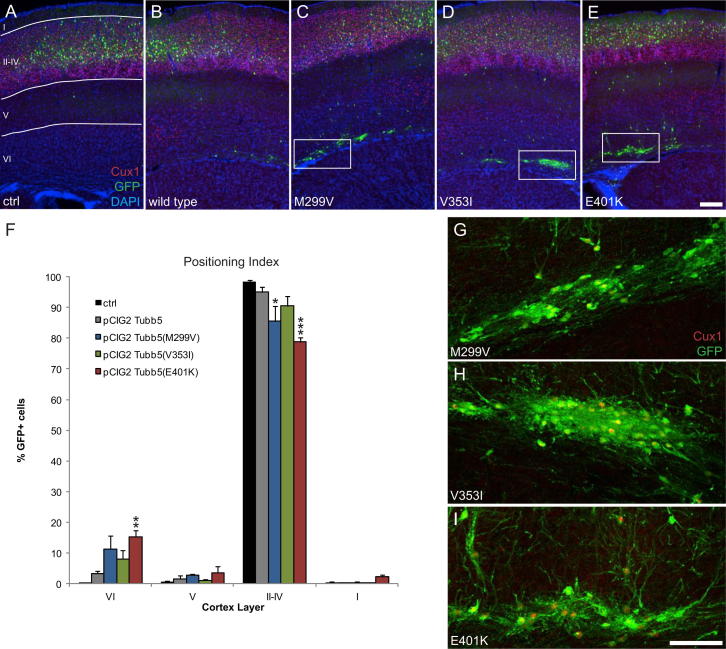
The formation of the mammalian cortex requires the generation, migration, and differentiation of neurons. The vital role that the microtubule cytoskeleton plays in these cellular processes is reflected by the discovery that mutations in various tubulin isotypes cause different neurodevelopmental diseases, including lissencephaly (TUBA1A), polymicrogyria (TUBA1A, TUBB2B, TUBB3), and an ocular motility disorder (TUBB3). Here, we show that Tubb5 is expressed in neurogenic progenitors in the mouse and that its depletion in vivo perturbs the cell cycle of progenitors and alters the position of migrating neurons. We report the occurrence of three microcephalic patients with structural brain abnormalities harboring de novo mutations in TUBB5 (M299V, V353I, and E401K). These mutant proteins, which affect the chaperone-dependent assembly of tubulin heterodimers in different ways, disrupt neurogenic division and/or migration in vivo. Our results provide insight into the functional repertoire of the tubulin gene family, specifically implicating TUBB5 in embryonic neurogenesis and microcephaly.
Copyright © 2012 The Authors. Published by Elsevier Inc. All rights reserved.
Figures


Similar articles
-
Mutations in the murine homologue of TUBB5 cause microcephaly by perturbing cell cycle progression and inducing p53-associated apoptosis.Development. 2016 Apr 1;143(7):1126-33. doi: 10.1242/dev.131516. Epub 2016 Feb 22. Development. 2016. PMID: 26903504
-
TUBA1A tubulinopathy mutants disrupt neuron morphogenesis and override XMAP215/Stu2 regulation of microtubule dynamics.Elife. 2022 May 5;11:e76189. doi: 10.7554/eLife.76189. Elife. 2022. PMID: 35511030 Free PMC article.
-
The wide spectrum of tubulinopathies: what are the key features for the diagnosis?Brain. 2014 Jun;137(Pt 6):1676-700. doi: 10.1093/brain/awu082. Brain. 2014. PMID: 24860126
-
Insights on the Role of α- and β-Tubulin Isotypes in Early Brain Development.Mol Neurobiol. 2023 Jul;60(7):3803-3823. doi: 10.1007/s12035-023-03302-1. Epub 2023 Mar 21. Mol Neurobiol. 2023. PMID: 36943622 Review.
-
The mutational and phenotypic spectrum of TUBA1A-associated tubulinopathy.Orphanet J Rare Dis. 2019 Feb 11;14(1):38. doi: 10.1186/s13023-019-1020-x. Orphanet J Rare Dis. 2019. PMID: 30744660 Free PMC article. Review.
Cited by
-
SMIFH2 has effects on Formins and p53 that perturb the cell cytoskeleton.Sci Rep. 2015 Apr 30;5:9802. doi: 10.1038/srep09802. Sci Rep. 2015. PMID: 25925024 Free PMC article.
-
Aberrant proteins expressed in skin fibroblasts of Parkinson's disease patients carrying heterozygous variants of glucocerebrosidase and parkin genes.Biomed Rep. 2021 Apr;14(4):36. doi: 10.3892/br.2021.1412. Epub 2021 Feb 17. Biomed Rep. 2021. PMID: 33732455 Free PMC article.
-
Clinical and Functional Characterization of the Recurrent TUBA1A p.(Arg2His) Mutation.Brain Sci. 2018 Aug 7;8(8):145. doi: 10.3390/brainsci8080145. Brain Sci. 2018. PMID: 30087272 Free PMC article.
-
Ultra-high-field MR imaging in polymicrogyria and epilepsy.AJNR Am J Neuroradiol. 2015 Feb;36(2):309-16. doi: 10.3174/ajnr.A4116. Epub 2014 Sep 25. AJNR Am J Neuroradiol. 2015. PMID: 25258368 Free PMC article.
-
Mutations in TUBB8 and Human Oocyte Meiotic Arrest.N Engl J Med. 2016 Jan 21;374(3):223-32. doi: 10.1056/NEJMoa1510791. N Engl J Med. 2016. PMID: 26789871 Free PMC article.
References
-
- Ayala R., Shu T., Tsai L.H. Trekking across the brain: the journey of neuronal migration. Cell. 2007;128:29–43. - PubMed
-
- Cowan N.J., Lewis S.A. Type II chaperonins, prefoldin, and the tubulin-specific chaperones. Adv. Protein Chem. 2001;59:73–104. - PubMed
-
- Feng Y., Walsh C.A. Protein-protein interactions, cytoskeletal regulation and neuronal migration. Nat. Rev. Neurosci. 2001;2:408–416. - PubMed
Supplemental References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
