

Coactivator-associated arginine methyltransferase 1 regulates fetal hematopoiesis and thymocyte development
- PMID: 23248263
- PMCID: PMC3538901
- DOI: 10.4049/jimmunol.1102513
Coactivator-associated arginine methyltransferase 1 regulates fetal hematopoiesis and thymocyte development
Abstract
Coactivator-associated arginine methyltransferase 1 (CARM1) is a protein arginine methyltransferase that methylates histones and transcriptional regulators. We previously reported that the absence of CARM1 partially blocks thymocyte differentiation at embryonic day 18.5 (E18.5). In this study, we find that reduced thymopoiesis in Carm1(-/-) mice is due to a defect in the fetal hematopoietic compartment rather than in the thymic stroma. To determine the cellular basis for impaired thymopoiesis, we examined the number and function of fetal liver (FL) and bone marrow cells. Despite markedly reduced cellularity of hematopoietic progenitors in E18.5 bone marrow, the number of long-term hematopoietic stem cells and downstream subsets was not reduced in Carm1(-/-) E14.5 or E18.5 FL. Nevertheless, competitive reconstitution assays revealed a deficit in the ability of Carm1(-/-) FL cells to contribute to hematopoiesis. Furthermore, impaired differentiation of Carm1(-/-) FL cells in a CARM1-sufficient host showed that CARM1 is required cell autonomously in hematopoietic cells. Coculture of Carm1(-/-) FL cells on OP9-DL1 monolayers showed that CARM1 is required for survival of hematopoietic progenitors under conditions that promote differentiation. Taken together, this report demonstrates that CARM1 is a key epigenetic regulator of hematopoiesis that affects multiple lineages at various stages of differentiation.
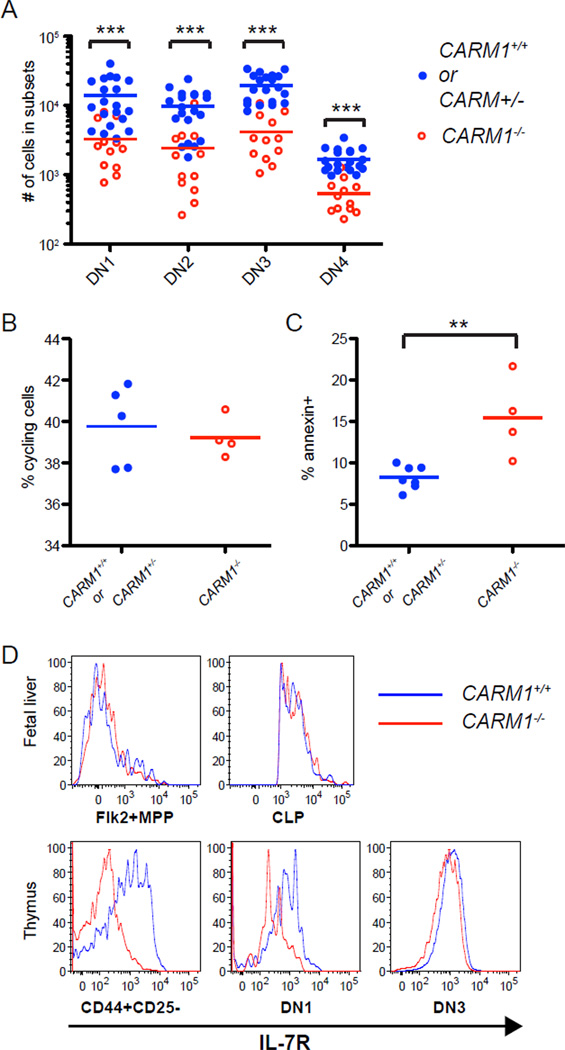
Figures


Similar articles
-
Loss of CARM1 results in hypomethylation of thymocyte cyclic AMP-regulated phosphoprotein and deregulated early T cell development.J Biol Chem. 2004 Jun 11;279(24):25339-44. doi: 10.1074/jbc.M402544200. Epub 2004 Apr 19. J Biol Chem. 2004. PMID: 15096520
-
CARM1 (PRMT4) Acts as a Transcriptional Coactivator during Retinoic Acid-Induced Embryonic Stem Cell Differentiation.J Mol Biol. 2018 Oct 19;430(21):4168-4182. doi: 10.1016/j.jmb.2018.08.014. Epub 2018 Aug 25. J Mol Biol. 2018. PMID: 30153436 Free PMC article.
-
Enzymatic activity is required for the in vivo functions of CARM1.J Biol Chem. 2010 Jan 8;285(2):1147-52. doi: 10.1074/jbc.M109.035865. Epub 2009 Nov 5. J Biol Chem. 2010. PMID: 19897492 Free PMC article.
-
Cyclical mobilization and gated importation of thymocyte progenitors in the adult mouse: evidence for a thymus-bone marrow feedback loop.Immunol Rev. 2006 Feb;209:58-75. doi: 10.1111/j.0105-2896.2006.00354.x. Immunol Rev. 2006. PMID: 16448534 Review.
-
Differential requirements for Wnt and Notch signaling in hematopoietic versus thymic niches.Ann N Y Acad Sci. 2012 Aug;1266:78-93. doi: 10.1111/j.1749-6632.2012.06626.x. Ann N Y Acad Sci. 2012. PMID: 22901260 Review.
Cited by
-
Coactivator-associated arginine methyltransferase 1: A versatile player in cell differentiation and development.Genes Dis. 2022 Jun 3;10(6):2383-2392. doi: 10.1016/j.gendis.2022.05.021. eCollection 2023 Nov. Genes Dis. 2022. PMID: 37554200 Free PMC article. Review.
-
Arginine Methylation by PRMT1 Regulates Muscle Stem Cell Fate.Mol Cell Biol. 2017 Jan 19;37(3):e00457-16. doi: 10.1128/MCB.00457-16. Print 2017 Feb 1. Mol Cell Biol. 2017. PMID: 27849571 Free PMC article.
-
'Nomadic' Hematopoietic Stem Cells Navigate the Embryonic Landscape.Stem Cell Rev Rep. 2025 Apr;21(3):605-628. doi: 10.1007/s12015-025-10843-6. Epub 2025 Jan 9. Stem Cell Rev Rep. 2025. PMID: 39786676 Review.
-
A TR-FRET-based functional assay for screening activators of CARM1.Chembiochem. 2013 May 10;14(7):827-35. doi: 10.1002/cbic.201300029. Epub 2013 Apr 12. Chembiochem. 2013. PMID: 23585185 Free PMC article.
-
CARM1 accelerates the growth of liver cancer cells by enhancing ARAF.Genes Dis. 2024 Aug 23;12(1):101395. doi: 10.1016/j.gendis.2024.101395. eCollection 2025 Jan. Genes Dis. 2024. PMID: 39415943 Free PMC article. No abstract available.
References
-
- Chao MP, Seita J, Weissman IL. Establishment of a normal hematopoietic and leukemia stem cell hierarchy. Cold Spring Harb Symp Quant Biol. 2008;73:439–449. - PubMed
-
- Mohn F, Schubeler D. Genetics and epigenetics: stability and plasticity during cellular differentiation. Trends Genet. 2009;25:129–136. - PubMed
-
- Shafa M, Krawetz R, Rancourt DE. Returning to the stem state: epigenetics of recapitulating pre-differentiation chromatin structure. Bioessays. 2010;32:791–799. - PubMed
-
- Ji H, Ehrlich LI, Seita J, Murakami P, Doi A, Lindau P, Lee H, Aryee MJ, Irizarry RA, Kim K, Rossi DJ, Inlay MA, Serwold T, Karsunky H, Ho L, Daley GQ, Weissman IL, Feinberg AP. Comprehensive methylome map of lineage commitment from haematopoietic progenitors. Nature. 2010;467:338–342. - PMC - PubMed
-
- Parry RV, Ward SG. Protein arginine methylation: a new handle on T lymphocytes? Trends Immunol. 2010;31:164–169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
