

Cdc42-dependent structural development of auditory supporting cells is required for wound healing at adulthood
- PMID: 23248743
- PMCID: PMC3523287
- DOI: 10.1038/srep00978
Cdc42-dependent structural development of auditory supporting cells is required for wound healing at adulthood
Abstract
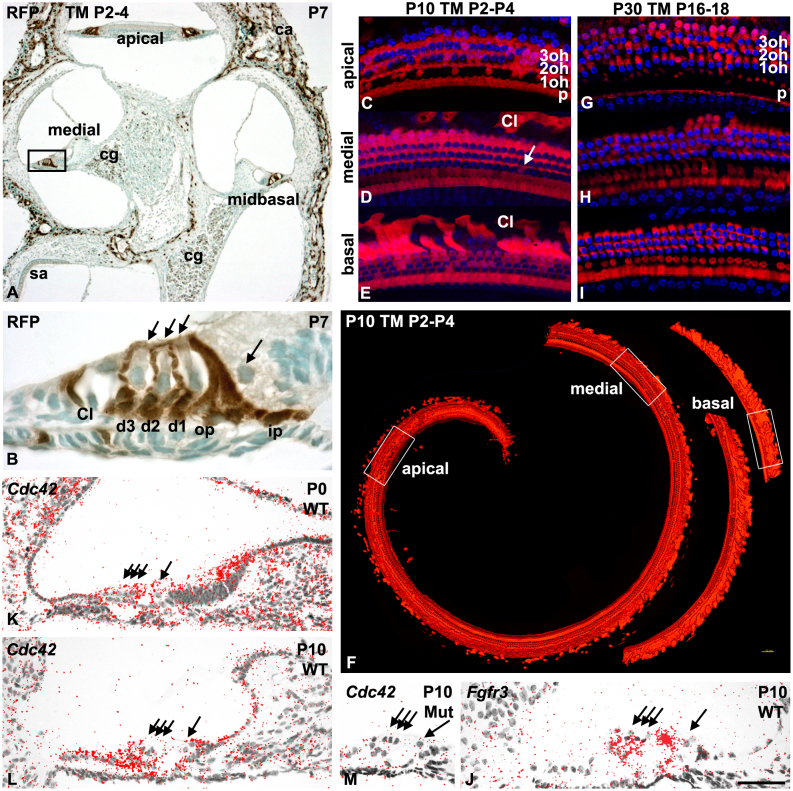
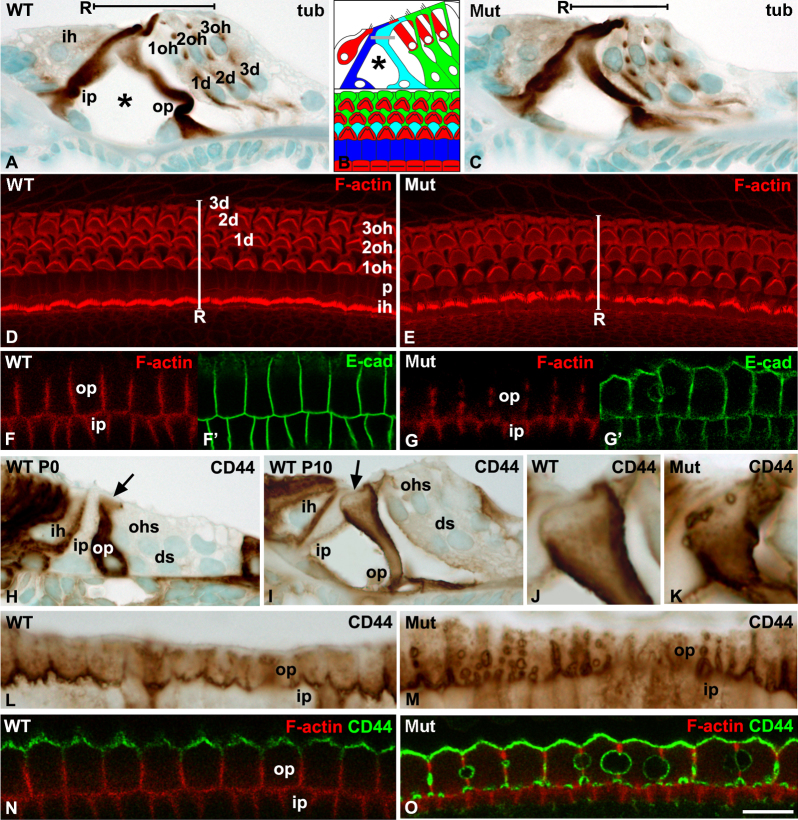
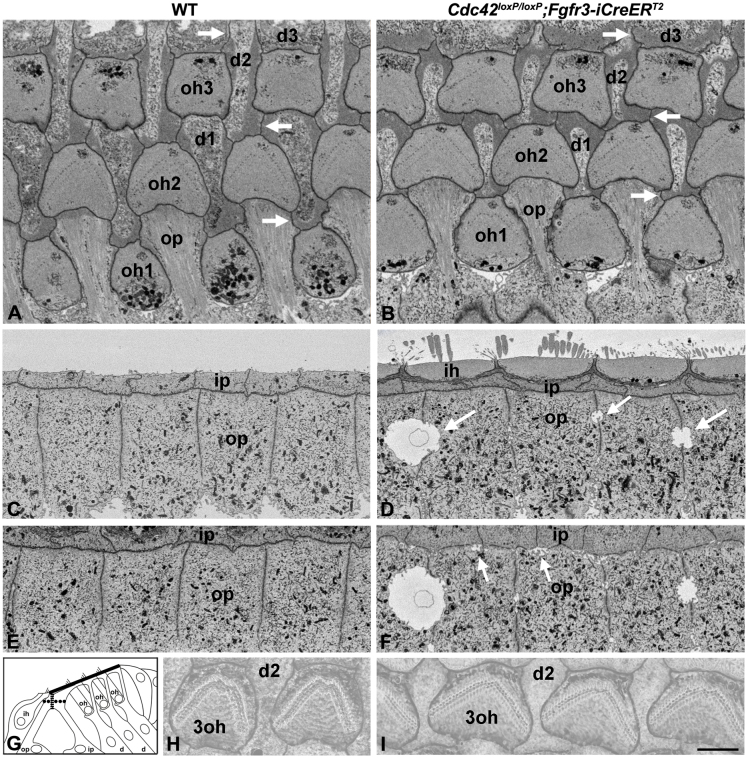
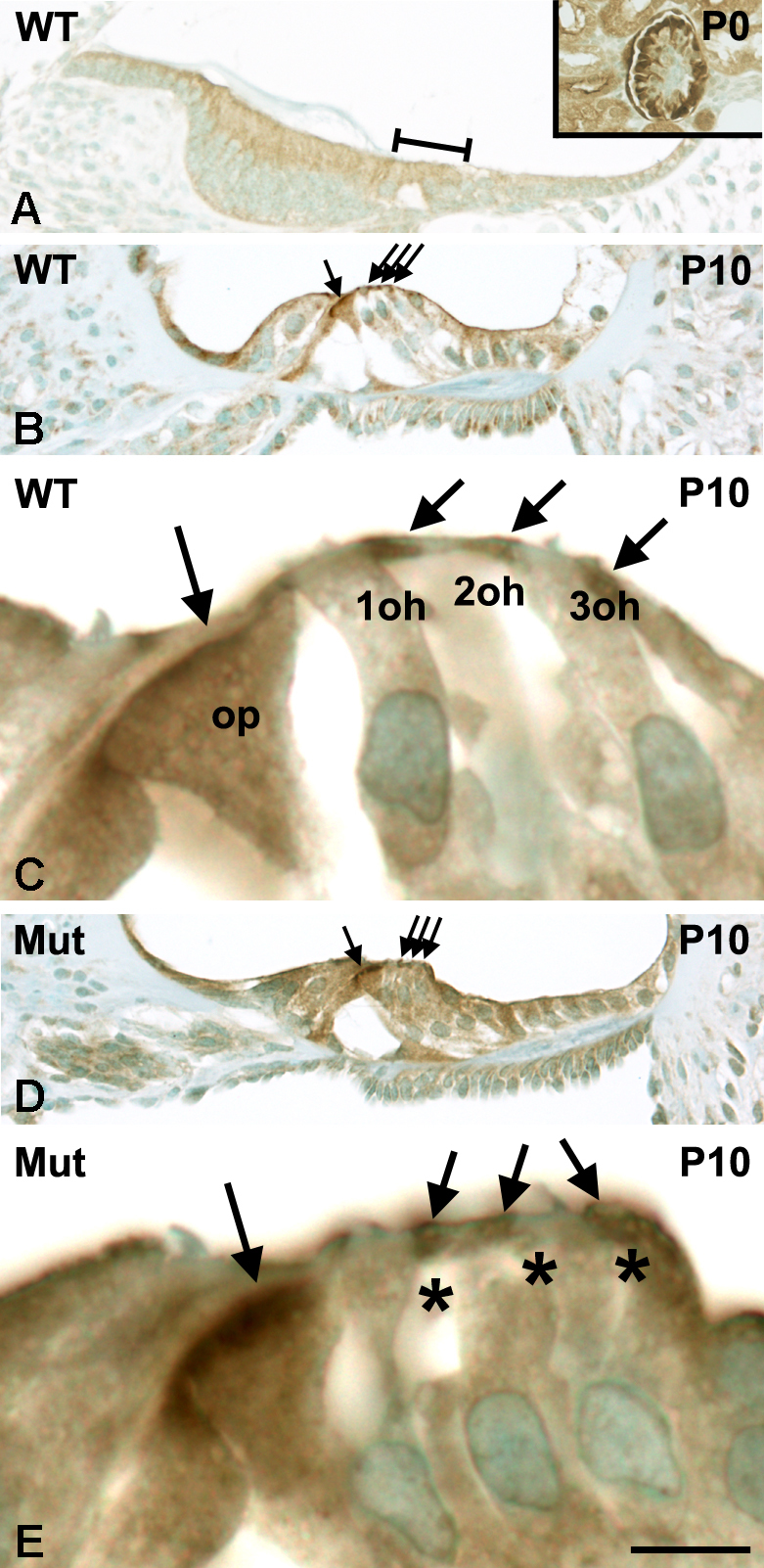
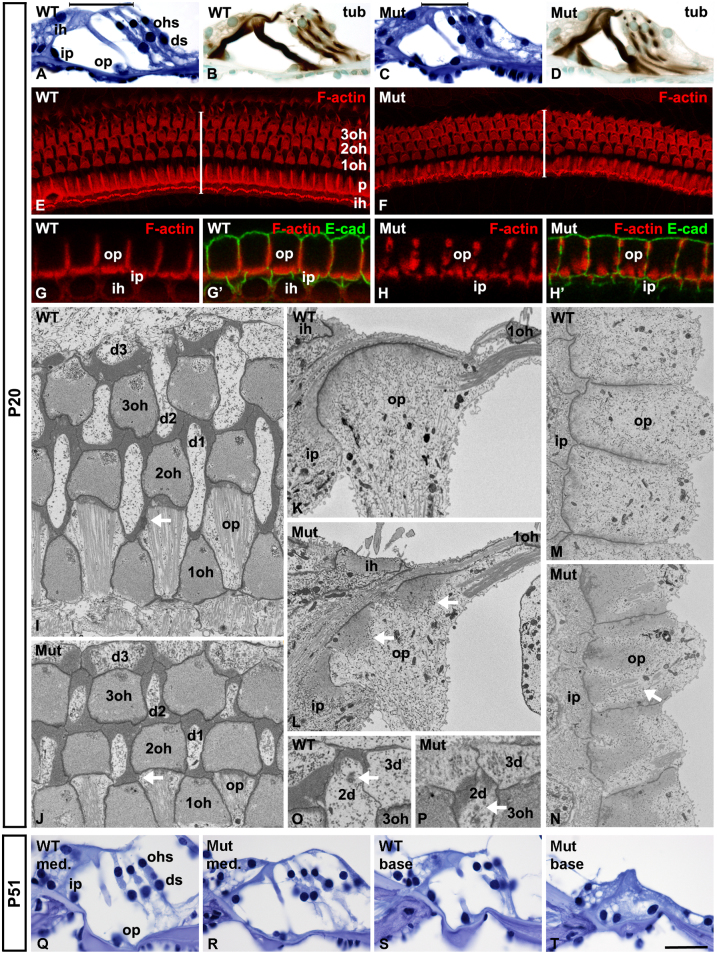
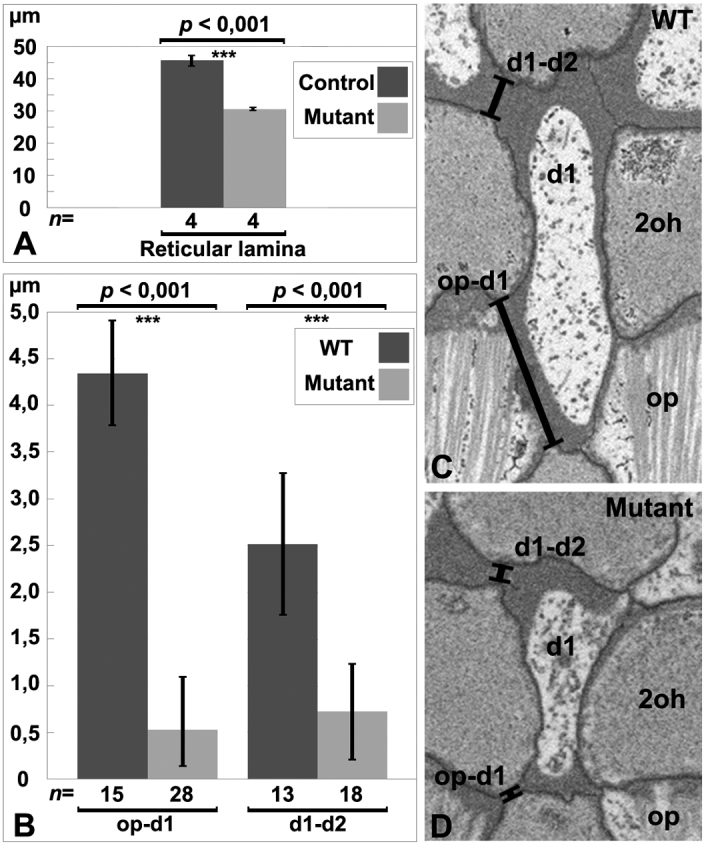
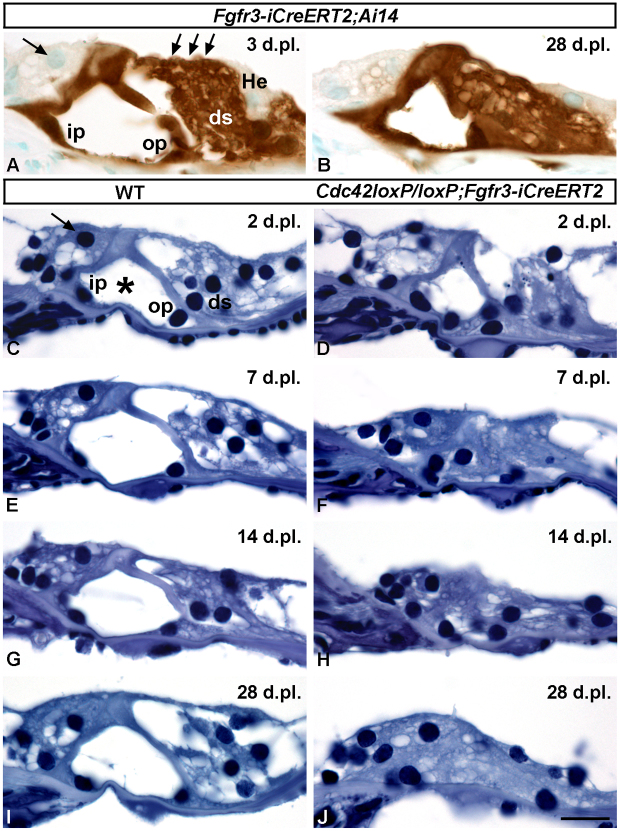
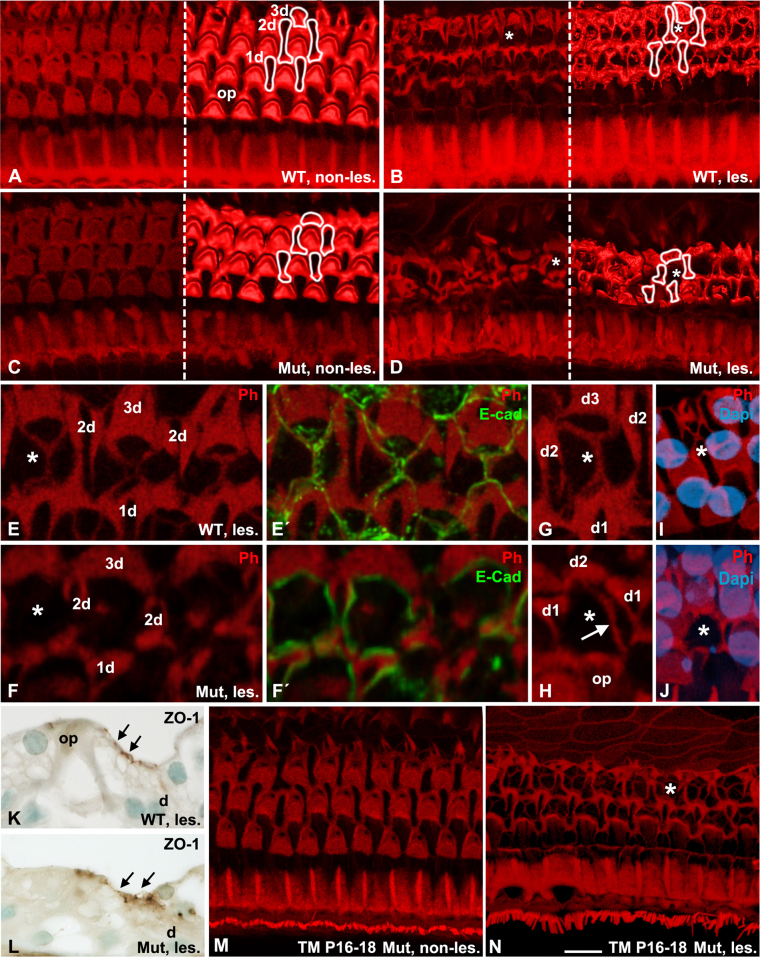
Cdc42 regulates the initial establishment of cytoskeletal and junctional structures, but only little is known about its role at later stages of cellular differentiation. We studied Cdc42's role in vivo in auditory supporting cells, epithelial cells with high structural complexity. Cdc42 inactivation was induced early postnatally using the Cdc42(loxP/loxP);Fgfr3-iCre-ER(T2) mice. Cdc42 depletion impaired elongation of adherens junctions and F-actin belts, leading to constriction of the sensory epithelial surface. Fragmented F-actin belts, junctions containing ectopic lumens and misexpression of a basolateral membrane protein in the apical domain were observed. These defects and changes in aPKCλ/ι expression suggested that apical polarization is impaired. Following a lesion at adulthood, supporting cells with Cdc42 loss-induced maturational defects collapsed and failed to remodel F-actin belts, a process that is critical to scar formation. Thus, Cdc42 is required for structural differentiation of auditory supporting cells and this proper maturation is necessary for wound healing in adults.
Figures
References
-
- Forge A. & Wright T. The molecular architecture of the inner ear. Br. Med. Bull 63, 5–24 (2002). - PubMed
-
- Raphael Y. & Altschuler R. A. Structure and innervation of the cochlea. Brain Res Bull 60, 397–422 (2003). - PubMed
-
- Colvin J. S., Bohne B. A., Harding G. W., McEwen D. G. & Ornitz D. M. Skeletal overgrowth and deafness in mice lacking fibroblast growth factor receptor 3. Nat. Genet 12, 390–397 (1996). - PubMed
-
- Gulley R. L. & Reese T. S. Intercellular junctions in the reticular lamina of the organ of Corti. J. Neurocytol 5, 479–507 (1976). - PubMed
-
- Leonova E. V. & Raphael Y. Organization of cell junctions and cytoskeleton in the reticular lamina in normal and ototoxically damaged organ of Corti. Hear. Res. 113, 14–28 (1997). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
