

Single-molecule nanometry for biological physics
- PMID: 23249673
- PMCID: PMC3549428
- DOI: 10.1088/0034-4885/76/1/016601
Single-molecule nanometry for biological physics
Abstract
Precision measurement is a hallmark of physics but the small length scale (∼nanometer) of elementary biological components and thermal fluctuations surrounding them challenge our ability to visualize their action. Here, we highlight the recent developments in single-molecule nanometry where the position of a single fluorescent molecule can be determined with nanometer precision, reaching the limit imposed by the shot noise, and the relative motion between two molecules can be determined with ∼0.3 nm precision at ∼1 ms time resolution, as well as how these new tools are providing fundamental insights into how motor proteins move on cellular highways. We will also discuss how interactions between three and four fluorescent molecules can be used to measure three and six coordinates, respectively, allowing us to correlate the movements of multiple components. Finally, we will discuss recent progress in combining angstrom-precision optical tweezers with single-molecule fluorescent detection, opening new windows for multi-dimensional single-molecule nanometry for biological physics.
Figures
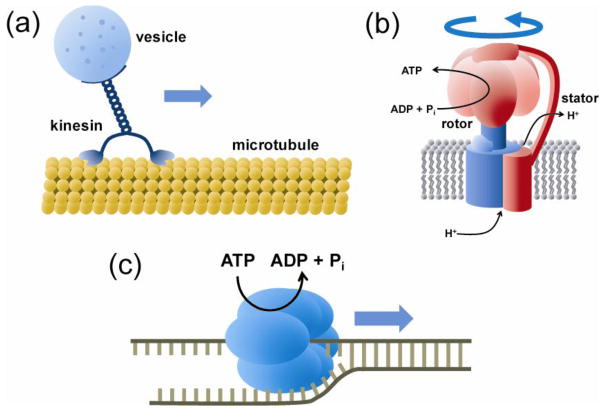
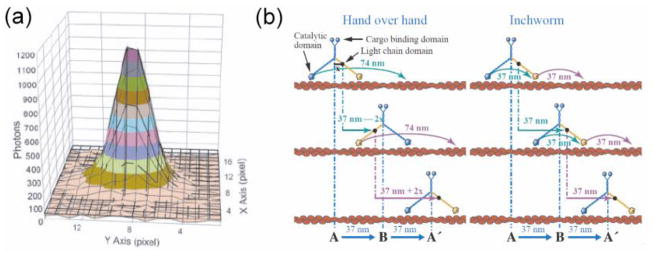
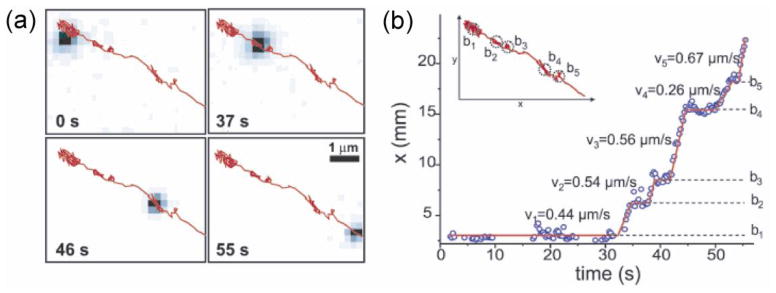
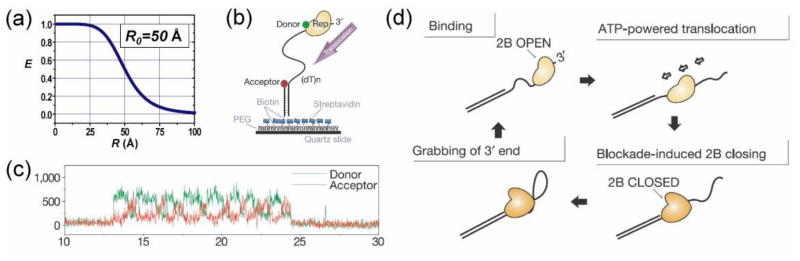
References
-
- Badali D, Gradinaru CC. The effect of Brownian motion of fluorescent probes on measuring nanoscale distances by F[o-umlaut]rster resonance energy transfer. The Journal of Chemical Physics. 2011;134:225102–11. - PubMed
-
- Bannai H, Levi S, Schweizer C, Dahan M, Triller A. Imaging the lateral diffusion of membrane molecules with quantum dots. Nat Protocols. 2007;1:2628–34. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources