

Neural-specific Sox2 input and differential Gli-binding affinity provide context and positional information in Shh-directed neural patterning
- PMID: 23249739
- PMCID: PMC3533082
- DOI: 10.1101/gad.207142.112
Neural-specific Sox2 input and differential Gli-binding affinity provide context and positional information in Shh-directed neural patterning
Abstract
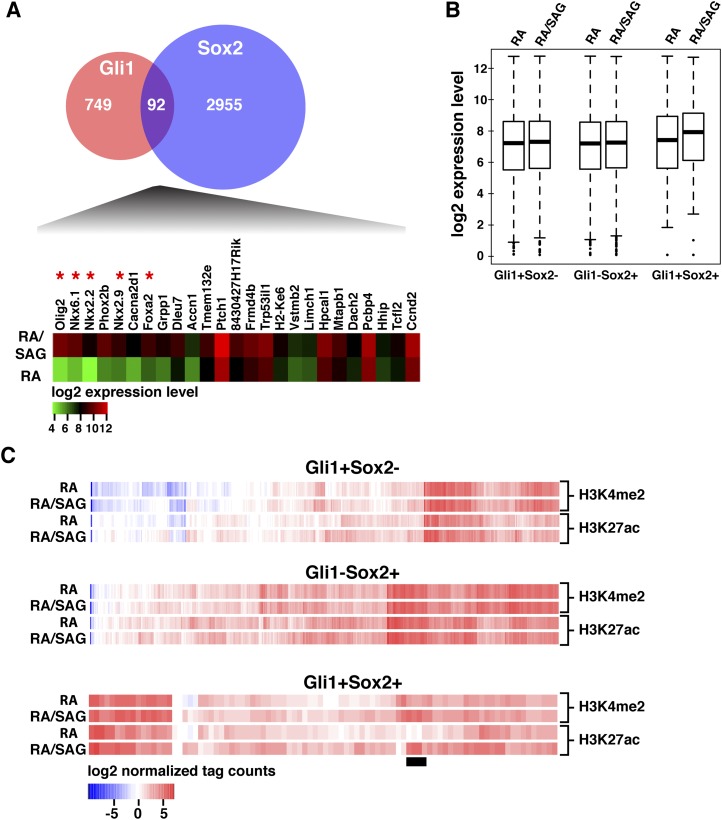
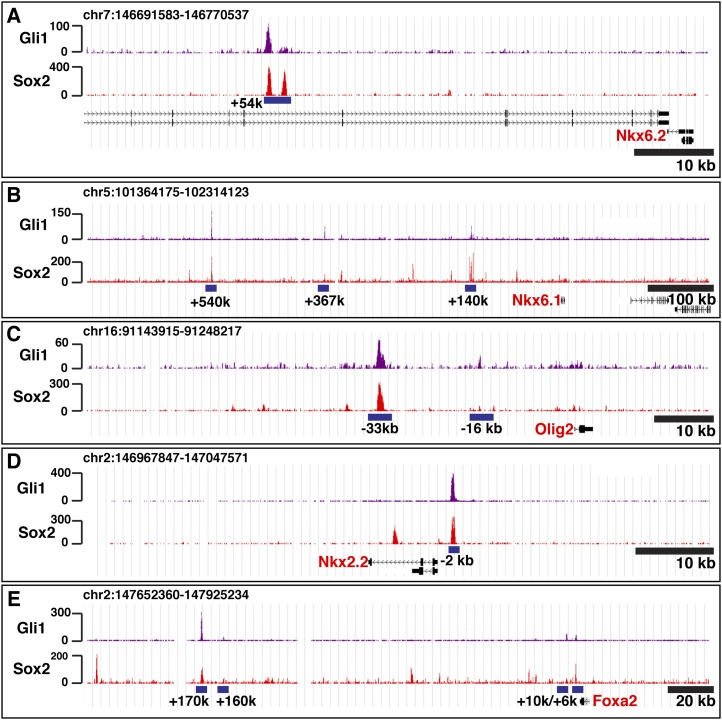
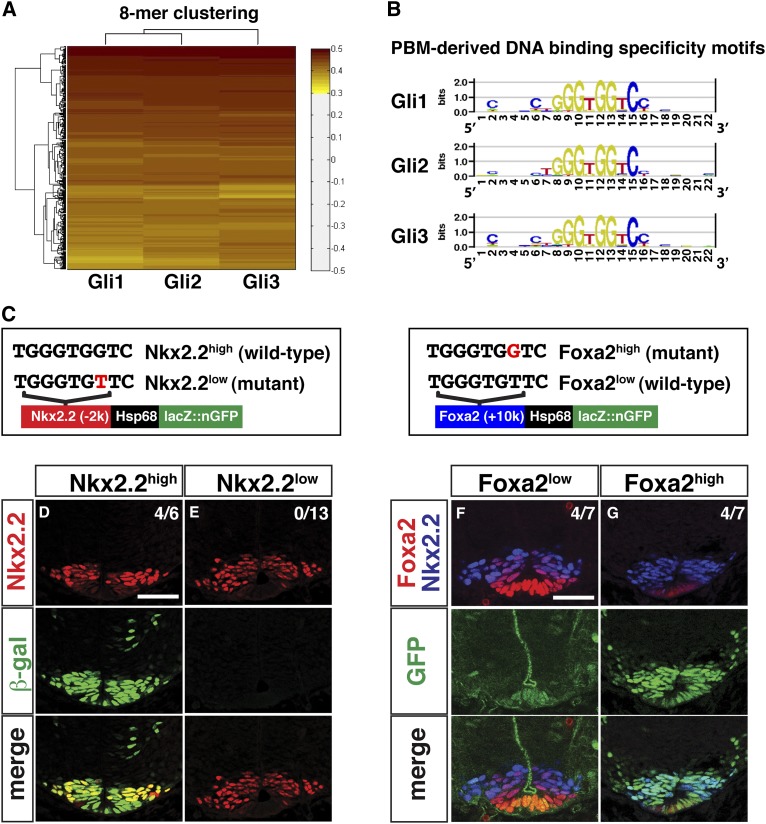
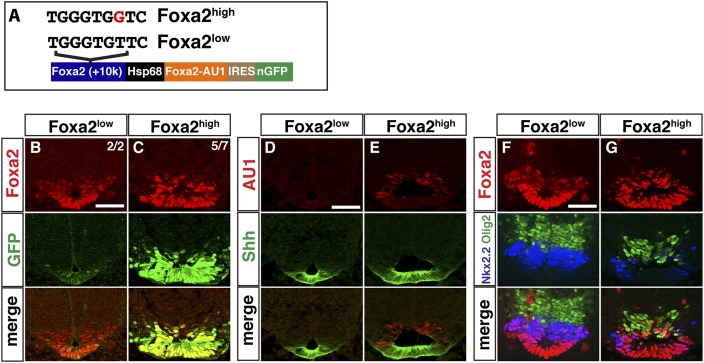
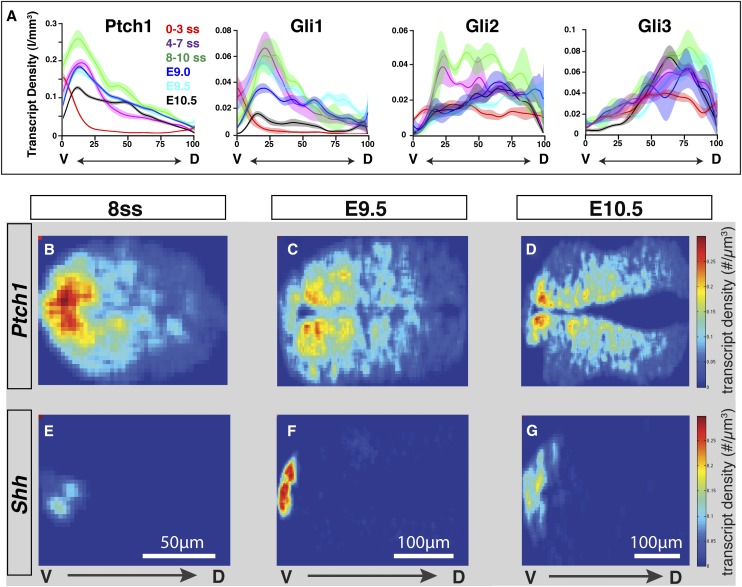
In the vertebrate neural tube, regional Sonic hedgehog (Shh) signaling invokes a time- and concentration-dependent induction of six different cell populations mediated through Gli transcriptional regulators. Elsewhere in the embryo, Shh/Gli responses invoke different tissue-appropriate regulatory programs. A genome-scale analysis of DNA binding by Gli1 and Sox2, a pan-neural determinant, identified a set of shared regulatory regions associated with key factors central to cell fate determination and neural tube patterning. Functional analysis in transgenic mice validates core enhancers for each of these factors and demonstrates the dual requirement for Gli1 and Sox2 inputs for neural enhancer activity. Furthermore, through an unbiased determination of Gli-binding site preferences and analysis of binding site variants in the developing mammalian CNS, we demonstrate that differential Gli-binding affinity underlies threshold-level activator responses to Shh input. In summary, our results highlight Sox2 input as a context-specific determinant of the neural-specific Shh response and differential Gli-binding site affinity as an important cis-regulatory property critical for interpreting Shh morphogen action in the mammalian neural tube.
Figures


Similar articles
-
A binding site for Gli proteins is essential for HNF-3beta floor plate enhancer activity in transgenics and can respond to Shh in vitro.Development. 1997 Apr;124(7):1313-22. doi: 10.1242/dev.124.7.1313. Development. 1997. PMID: 9118802
-
Mechanistic differences in the transcriptional interpretation of local and long-range Shh morphogen signaling.Dev Cell. 2012 Nov 13;23(5):1006-19. doi: 10.1016/j.devcel.2012.09.015. Dev Cell. 2012. PMID: 23153497
-
Ptch1 and Gli regulate Shh signalling dynamics via multiple mechanisms.Nat Commun. 2015 Apr 2;6:6709. doi: 10.1038/ncomms7709. Nat Commun. 2015. PMID: 25833741 Free PMC article.
-
The emergent design of the neural tube: prepattern, SHH morphogen and GLI code.Curr Opin Genet Dev. 2003 Oct;13(5):513-21. doi: 10.1016/j.gde.2003.08.005. Curr Opin Genet Dev. 2003. PMID: 14550418 Review.
-
Control of murine kidney development by sonic hedgehog and its GLI effectors.Cell Cycle. 2006 Jul;5(13):1426-30. doi: 10.4161/cc.5.13.2928. Epub 2006 Jul 1. Cell Cycle. 2006. PMID: 16855389 Review.
Cited by
-
HEDGEHOG/GLI-E2F1 axis modulates iASPP expression and function and regulates melanoma cell growth.Cell Death Differ. 2015 Dec;22(12):2006-19. doi: 10.1038/cdd.2015.56. Epub 2015 May 29. Cell Death Differ. 2015. PMID: 26024388 Free PMC article.
-
Hedgehog signaling is required for the maintenance of mesenchymal nephron progenitors.bioRxiv [Preprint]. 2024 May 5:2023.08.12.553098. doi: 10.1101/2023.08.12.553098. bioRxiv. 2024. PMID: 37645929 Free PMC article. Preprint.
-
Distinct regulatory mechanisms act to establish and maintain Pax3 expression in the developing neural tube.PLoS Genet. 2013;9(10):e1003811. doi: 10.1371/journal.pgen.1003811. Epub 2013 Oct 3. PLoS Genet. 2013. PMID: 24098141 Free PMC article.
-
Transcriptional regulation of graded Hedgehog signaling.Semin Cell Dev Biol. 2014 Sep;33:73-80. doi: 10.1016/j.semcdb.2014.05.010. Epub 2014 May 23. Semin Cell Dev Biol. 2014. PMID: 24862856 Free PMC article. Review.
-
The developmental hourglass model is applicable to the spinal cord based on single-cell transcriptomes and non-conserved cis-regulatory elements.Dev Growth Differ. 2021 Sep;63(7):372-391. doi: 10.1111/dgd.12750. Dev Growth Differ. 2021. PMID: 34473348 Free PMC article.
References
-
- Bai CB, Stephen D, Joyner AL 2004. All mouse ventral spinal cord patterning by hedgehog is Gli dependent and involves an activator function of Gli3. Dev Cell 6: 103–115 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases