

Th17 cells confer long-term adaptive immunity to oral mucosal Candida albicans infections
- PMID: 23250275
- PMCID: PMC3608691
- DOI: 10.1038/mi.2012.128
Th17 cells confer long-term adaptive immunity to oral mucosal Candida albicans infections
Abstract
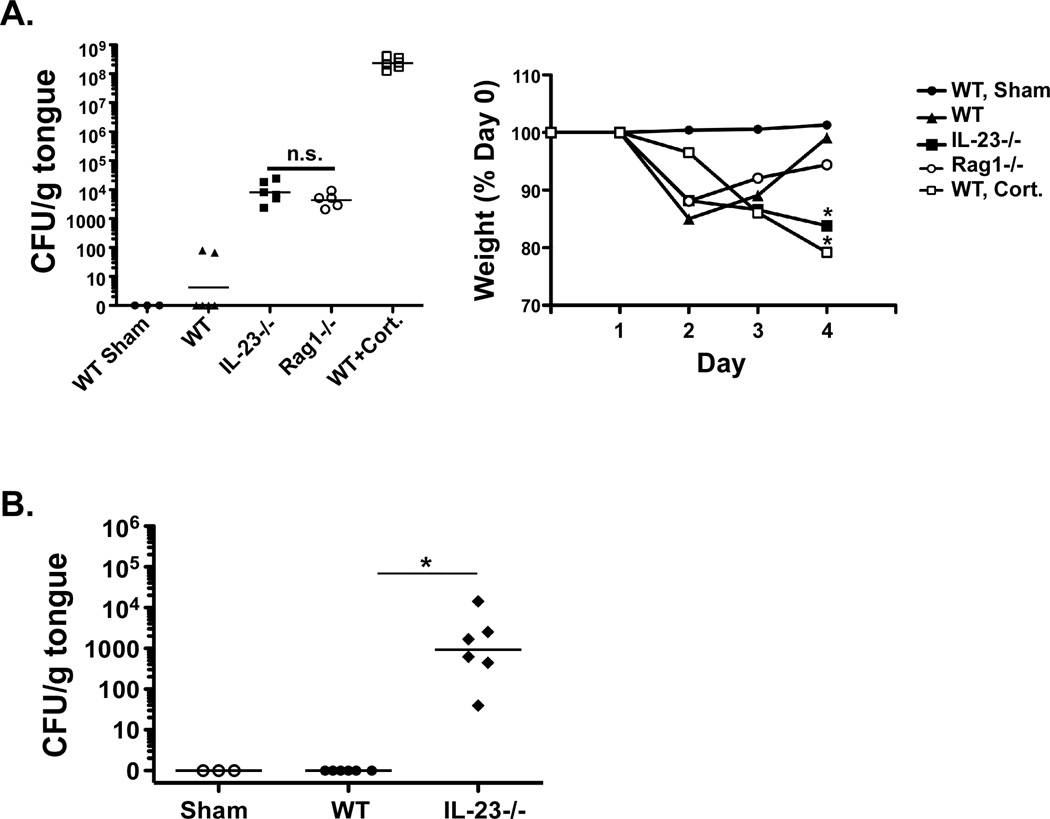
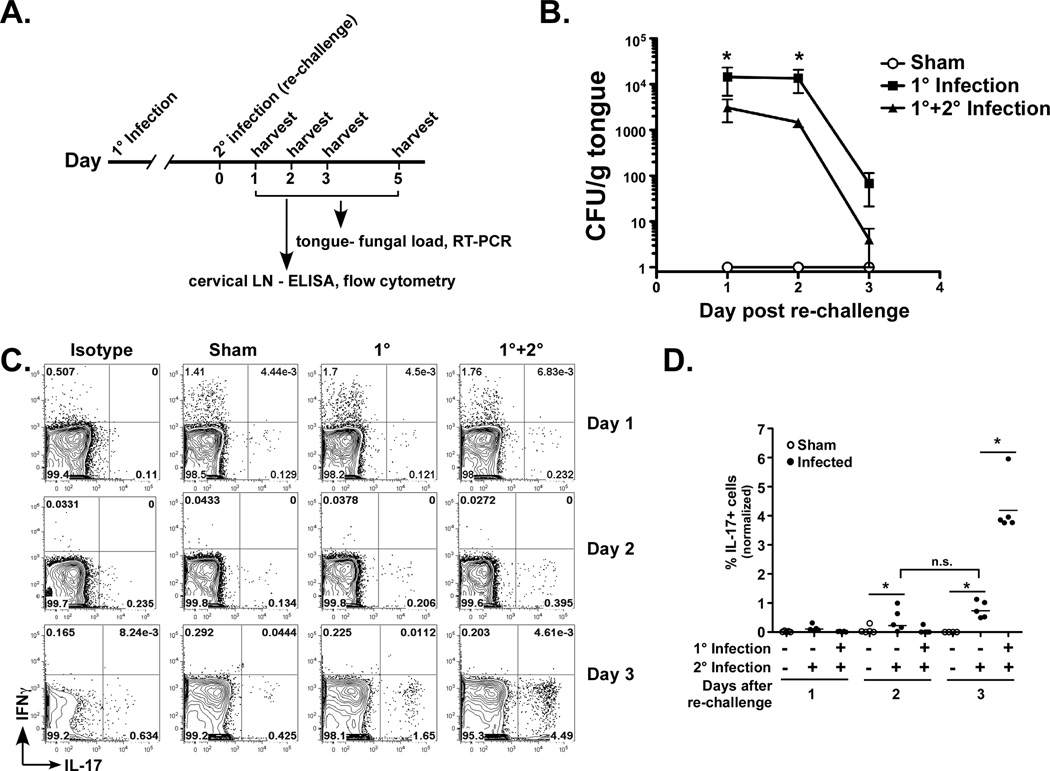
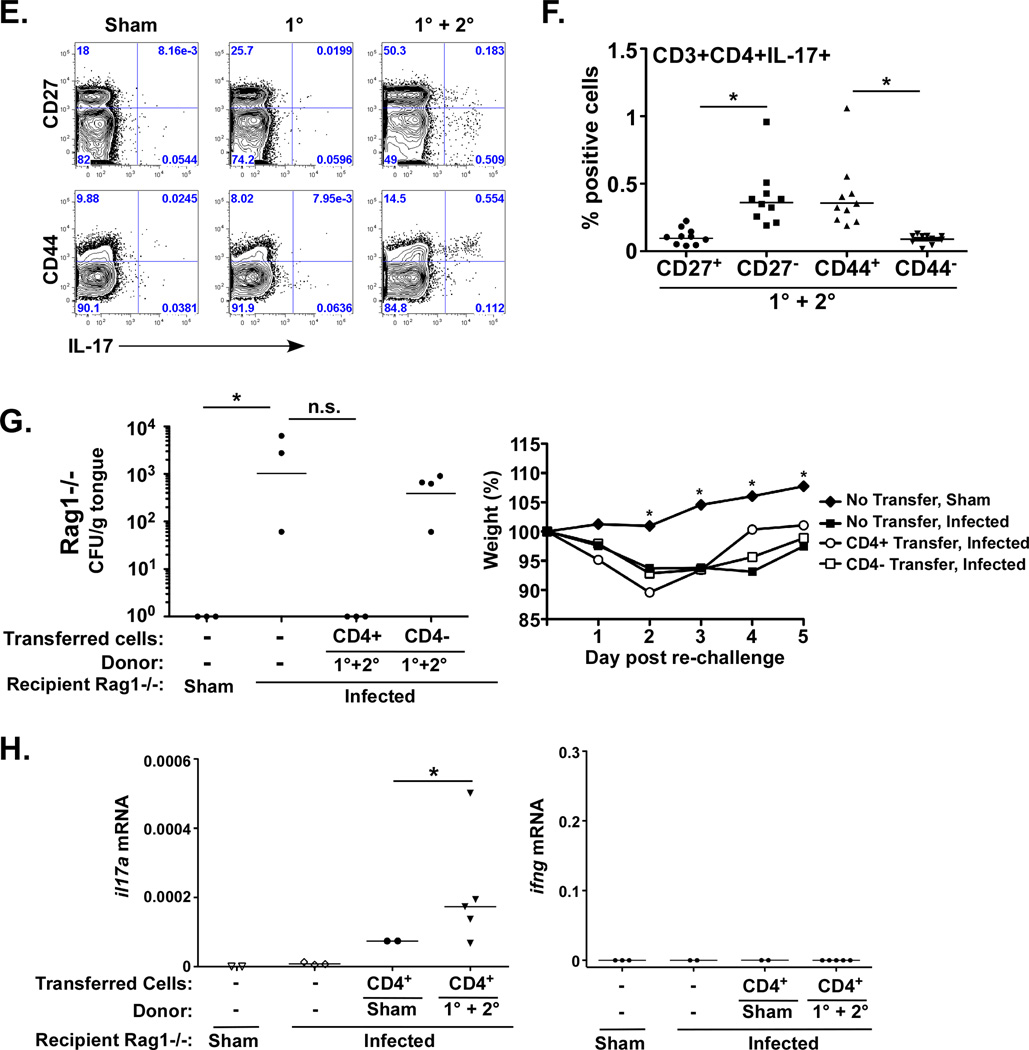
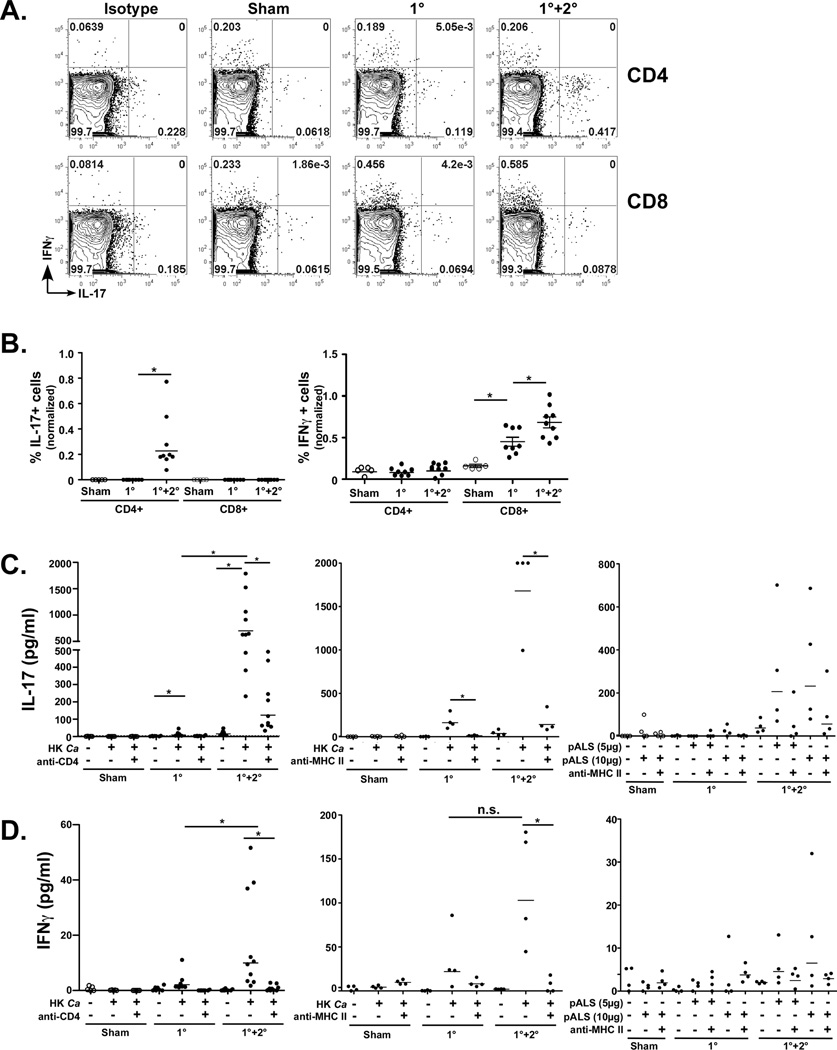
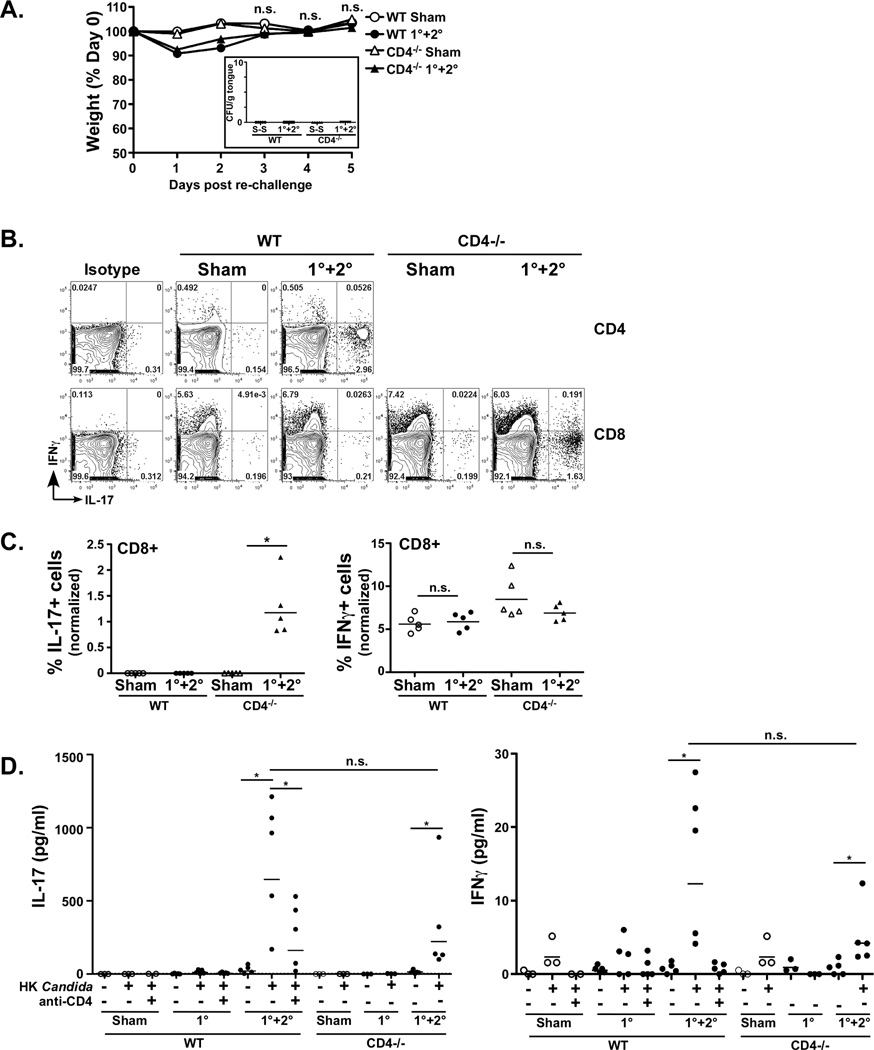
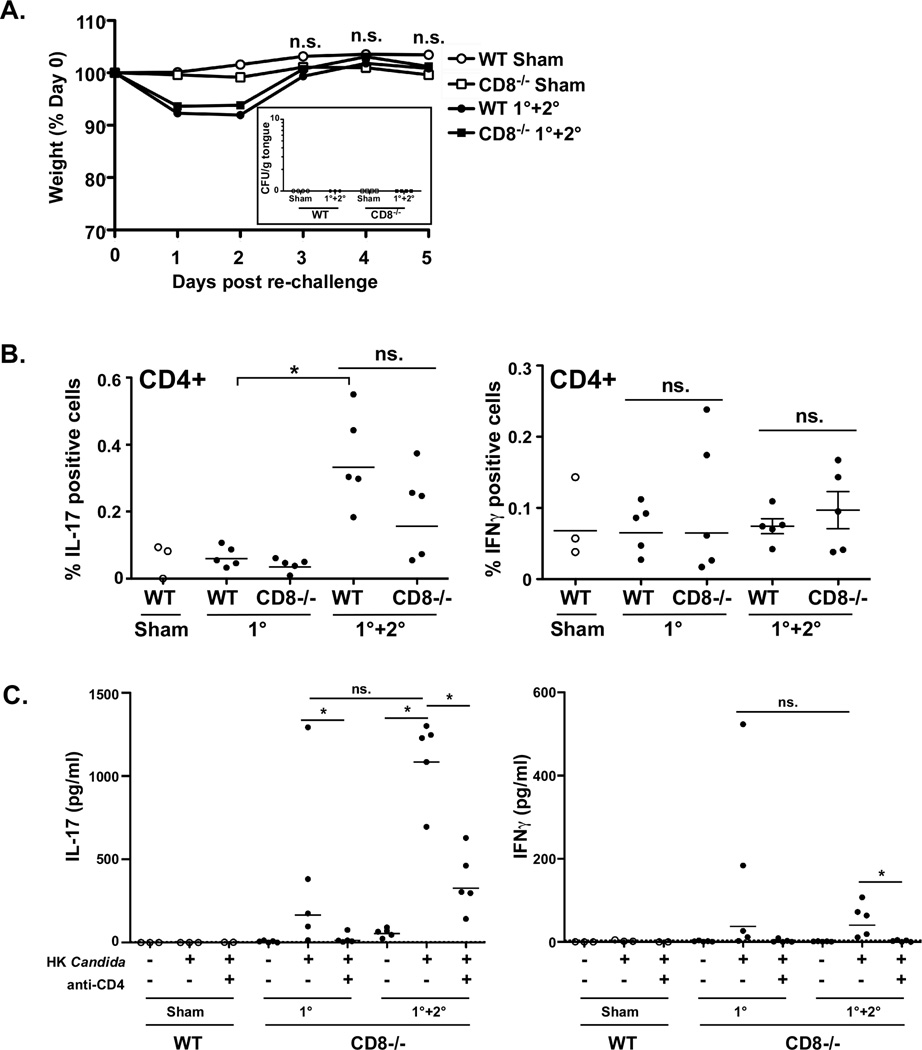
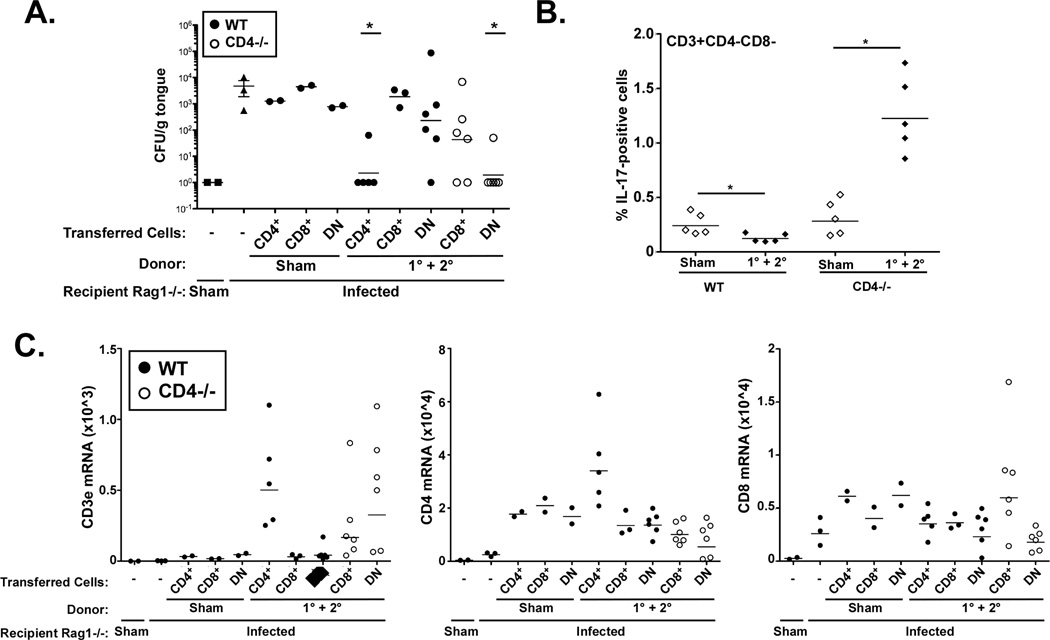
Oropharyngeal candidiasis (OPC) is an opportunistic infection caused by Candida albicans. Despite its prevalence, little is known about C. albicans-specific immunity in the oral mucosa. Vaccines against Candida generate both T helper type 1 (Th1) and Th17 responses, and considerable evidence implicates interleukin (IL)-17 in immunity to OPC. However, IL-17 is also produced by innate immune cells that are remarkably similar to Th17 cells, expressing the same markers and localizing to similar mucosal sites. To date, the relative contribution(s) of Th1, Th17, and innate IL-17-producing cells in OPC have not been clearly defined. Here, we sought to determine the nature and function of adaptive T-cell responses to OPC, using a new recall infection model. Mice subjected to infection and re-challenge with Candida mounted a robust and stable antigen-specific IL-17 response in CD4+ but not CD8+ T cells. There was little evidence for Th1 or Th1/Th17 responses. The Th17 response promoted accelerated fungal clearance, and Th17 cells could confer protection in Rag1-/- mice upon adoptive transfer. Surprisingly, CD4 deficiency did not cause OPC but was instead associated with compensatory IL-17 production by Tc17 and CD3+CD4-CD8- cells. Therefore, classic CD4+Th17 cells protect from OPC but can be compensated by other IL-17-producing cells in CD4-deficient hosts.
Conflict of interest statement
Conflicts of interest: SLG has received a research grant from Amgen, honoraria and travel reimbursements from Novartis, and consults for Lycera Corp.
Figures
Similar articles
-
The adaptor CARD9 is required for adaptive but not innate immunity to oral mucosal Candida albicans infections.Infect Immun. 2014 Mar;82(3):1173-80. doi: 10.1128/IAI.01335-13. Epub 2013 Dec 30. Infect Immun. 2014. PMID: 24379290 Free PMC article.
-
Oral-resident natural Th17 cells and γδ T cells control opportunistic Candida albicans infections.J Exp Med. 2014 Sep 22;211(10):2075-84. doi: 10.1084/jem.20130877. Epub 2014 Sep 8. J Exp Med. 2014. PMID: 25200028 Free PMC article.
-
Role of neutrophils in IL-17-dependent immunity to mucosal candidiasis.J Immunol. 2014 Feb 15;192(4):1745-52. doi: 10.4049/jimmunol.1302265. Epub 2014 Jan 17. J Immunol. 2014. PMID: 24442441 Free PMC article.
-
Immunity to Candida.Oral Dis. 2002;8 Suppl 2:69-75. doi: 10.1034/j.1601-0825.2002.00015.x. Oral Dis. 2002. PMID: 12164664 Review.
-
Host responses to Candida albicans: Th17 cells and mucosal candidiasis.Microbes Infect. 2010 Jul;12(7):518-27. doi: 10.1016/j.micinf.2010.03.013. Epub 2010 Apr 8. Microbes Infect. 2010. PMID: 20381638 Free PMC article. Review.
Cited by
-
IL-17-Mediated Immunity to the Opportunistic Fungal Pathogen Candida albicans.J Immunol. 2015 Aug 1;195(3):780-8. doi: 10.4049/jimmunol.1500909. J Immunol. 2015. PMID: 26188072 Free PMC article. Review.
-
Bio- and Nanotechnology as the Key for Clinical Application of Salivary Peptide Histatin: A Necessary Advance.Microorganisms. 2020 Jul 10;8(7):1024. doi: 10.3390/microorganisms8071024. Microorganisms. 2020. PMID: 32664360 Free PMC article. Review.
-
IκBζ is an essential mediator of immunity to oropharyngeal candidiasis.Cell Host Microbe. 2023 Oct 11;31(10):1700-1713.e4. doi: 10.1016/j.chom.2023.08.016. Epub 2023 Sep 18. Cell Host Microbe. 2023. PMID: 37725983 Free PMC article.
-
The dichotomous nature of T helper 17 cells.Nat Rev Immunol. 2017 Sep;17(9):535-544. doi: 10.1038/nri.2017.50. Epub 2017 May 30. Nat Rev Immunol. 2017. PMID: 28555673 Review.
-
MHC-II presentation by oral Langerhans cells impacts intraepithelial Tc17 abundance and Candida albicans oral infection via CD4 T cells.Front Oral Health. 2024 May 30;5:1408255. doi: 10.3389/froh.2024.1408255. eCollection 2024. Front Oral Health. 2024. PMID: 38872986 Free PMC article.
References
-
- Dongari-Bagtoglou A, Fidel P. The host cytokine responses and protective immunity in oropharyngeal candidiasis. J Dent Res. 2005;84:966–977. - PubMed
-
- Klein RS, Harris CA, Small CB, Moll B, Lesser M, Friedland GH. Oral candidiasis in high-risk patients as the initial manifestation of the acquired immunodeficiency syndrome. N Engl J Med. 1984 Aug 9;311(6):354–358. - PubMed
-
- Dong C. Diversification of T-helper-cell lineages: finding the family root of IL-17-producing cells. Nat Rev Immunol. 2006 Apr;6(4):329–333. - PubMed
-
- Steinman L. A brief history of T(H)17, the first major revision in the T(H)1/T(H)2 hypothesis of T cell-mediated tissue damage. Nature Med. 2007 Feb;13(2):139–145. - PubMed
-
- Farah C, Hu Y, Riminton S, Ashman R. Distinct roles for interleukin-12p40 and tumour necrosis factor in resistance to oral candidiasis defined by gene targeting. Oral Microbiol Immunol. 2006;21:252–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
