

Drosophila ryanodine receptors mediate general anesthesia by halothane
- PMID: 23254148
- PMCID: PMC3580016
- DOI: 10.1097/ALN.0b013e31827e52c6
Drosophila ryanodine receptors mediate general anesthesia by halothane
Abstract
Background: Although in vitro studies have identified numerous possible targets, the molecules that mediate the in vivo effects of volatile anesthetics remain largely unknown. The mammalian ryanodine receptor (Ryr) is a known halothane target, and the authors hypothesized that it has a central role in anesthesia.
Methods: Gene function of the Drosophila Ryr (dRyr) was manipulated in the whole body or in specific tissues using a collection of mutants and transgenes, and responses to halothane were measured with a reactive climbing assay. Cellular responses to halothane were studied using Ca imaging and patch clamp electrophysiology.
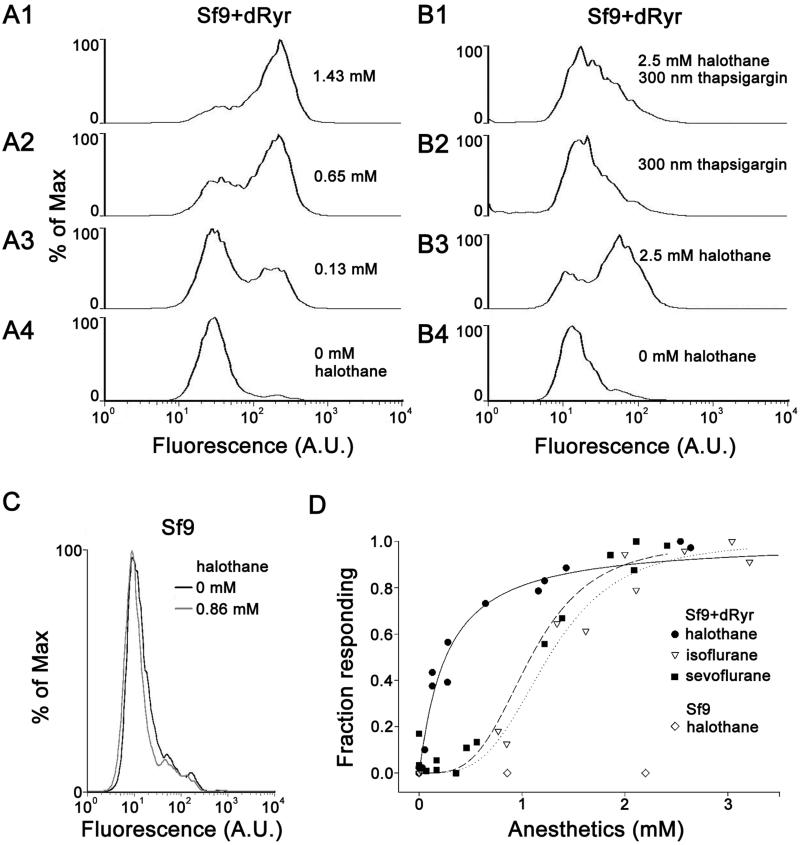
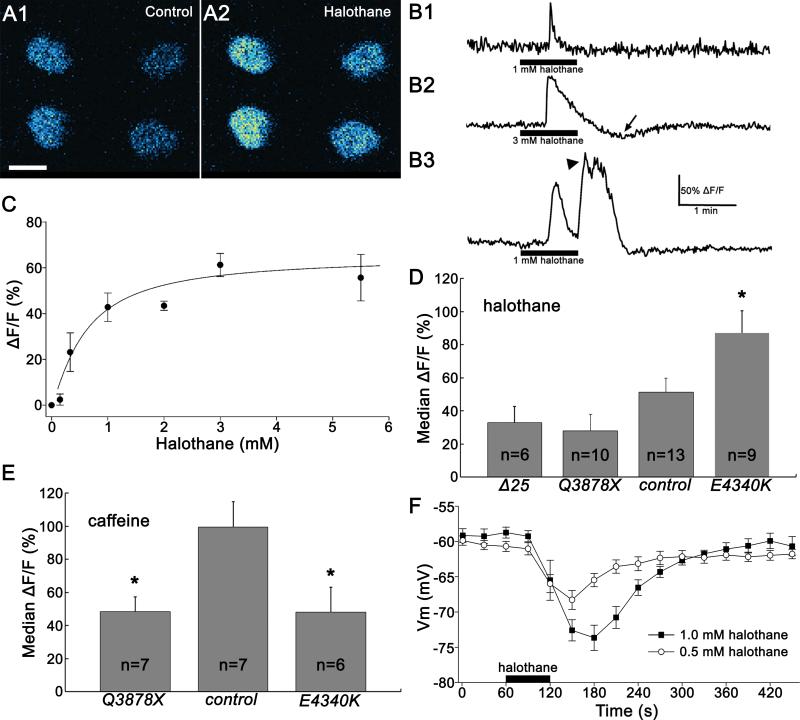
Results: Halothane potency strongly correlates with dRyr gene copy number, and missense mutations in regions known to be functionally important in the mammalian Ryrs gene cause dominant hypersensitivity. Tissue-specific manipulation of dRyr shows that expression in neurons and glia, but not muscle, mediates halothane sensitivity. In cultured cells, halothane-induced Ca efflux is strictly dRyr-dependent, suggesting a close interaction between halothane and dRyr. Ca imaging and electrophysiology of Drosophila central neurons reveal halothane-induced Ca flux that is altered in dRyr mutants and correlates with strong hyperpolarization.
Conclusions: In Drosophila, neurally expressed dRyr mediates a substantial proportion of the anesthetic effects of halothane in vivo, is potently activated by halothane in vitro, and activates an inhibitory conductance. The authors' results provide support for Ryr as an important mediator of immobilization by volatile anesthetics.
Figures
Similar articles
-
Selected contribution: effect of volatile anesthetics on cADP-ribose-induced Ca(2+) release system.J Appl Physiol (1985). 2001 Jul;91(1):516-21; discussion 504-5. doi: 10.1152/jappl.2001.91.1.516. J Appl Physiol (1985). 2001. PMID: 11408471
-
Voltage modulates halothane-triggered Ca2+ release in malignant hyperthermia-susceptible muscle.J Gen Physiol. 2018 Jan 2;150(1):111-125. doi: 10.1085/jgp.201711864. Epub 2017 Dec 15. J Gen Physiol. 2018. PMID: 29247050 Free PMC article.
-
Halothane-induced intracellular calcium release in cholinergic cells.Brain Res. 2001 Dec 7;921(1-2):106-14. doi: 10.1016/s0006-8993(01)03098-0. Brain Res. 2001. PMID: 11720716
-
Molecular cloning of cDNA encoding a drosophila ryanodine receptor and functional studies of the carboxyl-terminal calcium release channel.Biophys J. 2000 Mar;78(3):1270-81. doi: 10.1016/S0006-3495(00)76683-5. Biophys J. 2000. PMID: 10692315 Free PMC article.
-
Volatile general anesthetics produce hyperpolarization of Aplysia neurons by activation of a discrete population of baseline potassium channels.Anesthesiology. 1996 Oct;85(4):889-900. doi: 10.1097/00000542-199610000-00026. Anesthesiology. 1996. PMID: 8873561
Cited by
-
Neuronal processing of noxious thermal stimuli mediated by dendritic Ca(2+) influx in Drosophila somatosensory neurons.Elife. 2016 Feb 15;5:e12959. doi: 10.7554/eLife.12959. Elife. 2016. PMID: 26880554 Free PMC article.
-
Regulation of longevity by depolarization-induced activation of PLC-β-IP3R signaling in neurons.Proc Natl Acad Sci U S A. 2021 Apr 20;118(16):e2004253118. doi: 10.1073/pnas.2004253118. Proc Natl Acad Sci U S A. 2021. PMID: 33859040 Free PMC article.
-
Modality specific roles for metabotropic GABAergic signaling and calcium induced calcium release mechanisms in regulating cold nociception.Front Mol Neurosci. 2022 Sep 9;15:942548. doi: 10.3389/fnmol.2022.942548. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36157080 Free PMC article.
-
Extension of Health Span and Life Span in Drosophila by S107 Requires the calstabin Homologue FK506-BP2.J Biol Chem. 2016 Dec 9;291(50):26045-26055. doi: 10.1074/jbc.M116.758839. Epub 2016 Nov 1. J Biol Chem. 2016. PMID: 27803160 Free PMC article.
-
Isoflurane activates the type 1 ryanodine receptor to induce anesthesia in mice.PLoS Biol. 2025 Jun 3;23(6):e3003172. doi: 10.1371/journal.pbio.3003172. eCollection 2025 Jun. PLoS Biol. 2025. PMID: 40460053 Free PMC article.
References
-
- Rudolph U, Antkowiak B. Molecular and neuronal substrates for general anaesthetics. Nature Rev Neurosci. 2004;5:709–20. - PubMed
-
- Humphrey JA, Hamming KS, Thacker CM, Scott RL, Sedensky MM, Snutch TP, Morgan PG, Nash HA. A putative cation channel and its novel regulator: Cross-species conservation of effects on general anesthesia. Curr Biol. 2007;17:624–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
