

Insights into bilaterian evolution from three spiralian genomes
- PMID: 23254933
- PMCID: PMC4085046
- DOI: 10.1038/nature11696
Insights into bilaterian evolution from three spiralian genomes
Abstract
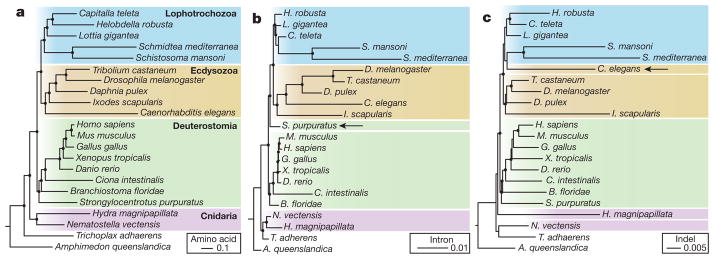
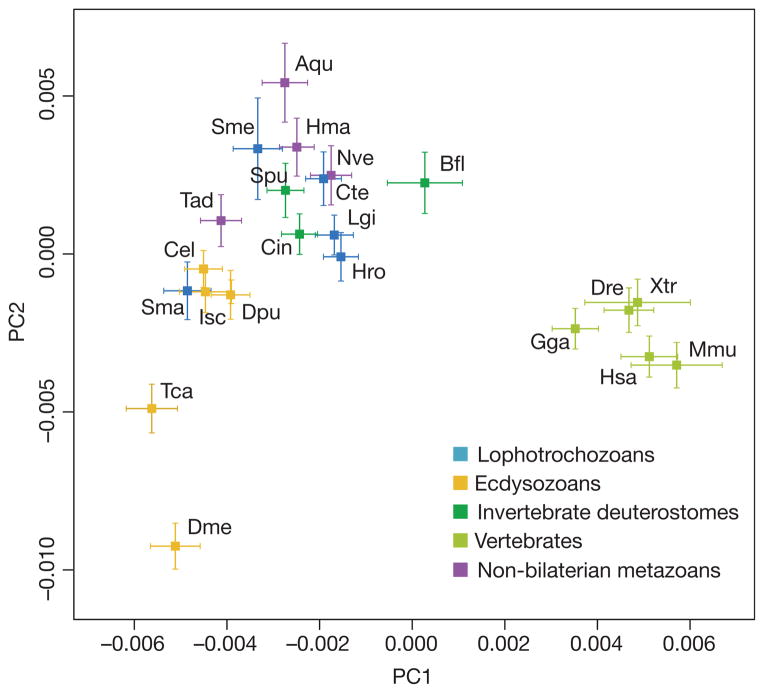
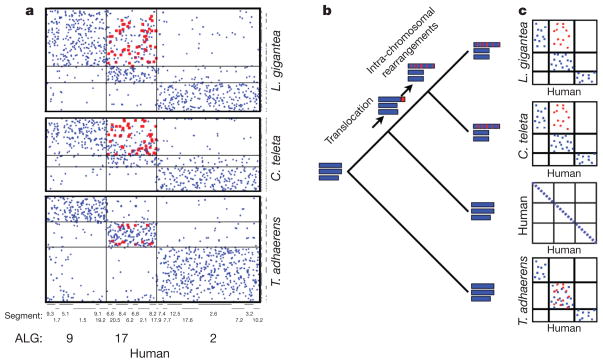
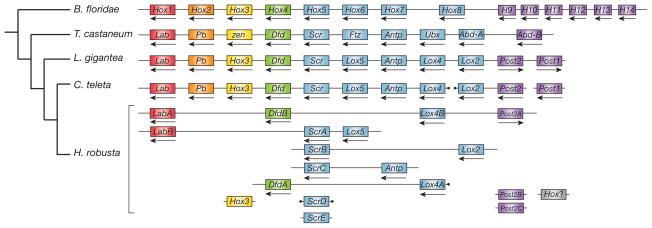
Current genomic perspectives on animal diversity neglect two prominent phyla, the molluscs and annelids, that together account for nearly one-third of known marine species and are important both ecologically and as experimental systems in classical embryology. Here we describe the draft genomes of the owl limpet (Lottia gigantea), a marine polychaete (Capitella teleta) and a freshwater leech (Helobdella robusta), and compare them with other animal genomes to investigate the origin and diversification of bilaterians from a genomic perspective. We find that the genome organization, gene structure and functional content of these species are more similar to those of some invertebrate deuterostome genomes (for example, amphioxus and sea urchin) than those of other protostomes that have been sequenced to date (flies, nematodes and flatworms). The conservation of these genomic features enables us to expand the inventory of genes present in the last common bilaterian ancestor, establish the tripartite diversification of bilaterians using multiple genomic characteristics and identify ancient conserved long- and short-range genetic linkages across metazoans. Superimposed on this broadly conserved pan-bilaterian background we find examples of lineage-specific genome evolution, including varying rates of rearrangement, intron gain and loss, expansions and contractions of gene families, and the evolution of clade-specific genes that produce the unique content of each genome.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Wilson EB. The cell-lineage of Nereis. A contribution tothe cytogeny of the annelid body. J Morphol. 1892;6:361–480.
-
- Conklin EG. The Embryology of Crepidula: a Contribution to the Cell Lineage and Early Development of Some Marine Gasteropods. Ginn & Company; 1897.
-
- Henry JQ, Hejnol A, Perry KJ, Martindale MQ. Homology of ciliary bands in spiralian trochophores. Integr Comp Biol. 2007;47:865–871. - PubMed
-
- Fedonkin MA, Waggoner BM. The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism. Nature. 1997;388:868–871.
-
- Maloof AC, et al. The earliest Cambrian record of animals and ocean geochemical change. Geol Soc Am Bull. 2010;122:1731–1774.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
