

Generating super-shedders: co-infection increases bacterial load and egg production of a gastrointestinal helminth
- PMID: 23256186
- PMCID: PMC3565725
- DOI: 10.1098/rsif.2012.0588
Generating super-shedders: co-infection increases bacterial load and egg production of a gastrointestinal helminth
Abstract
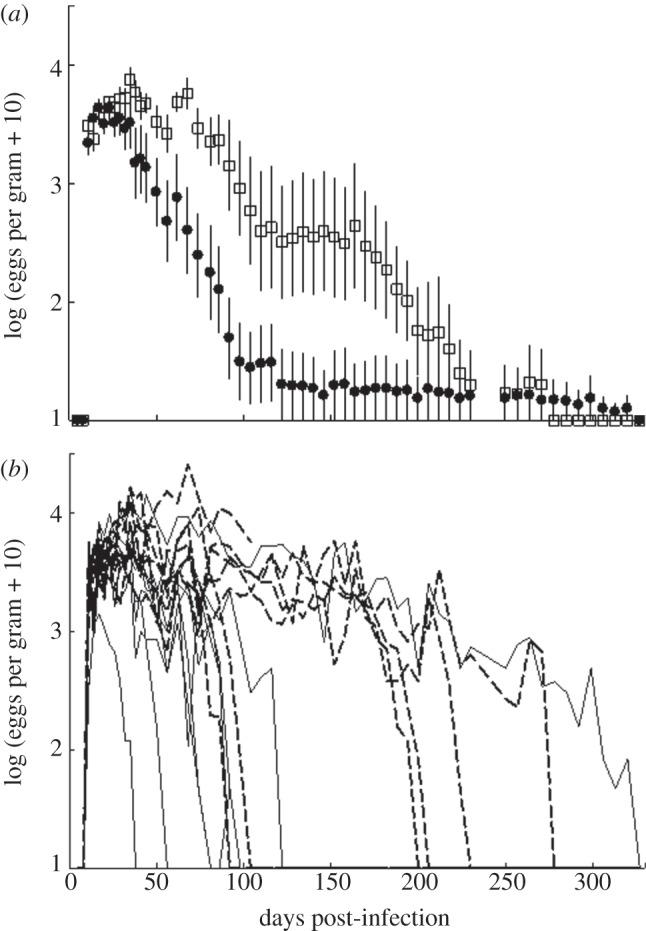
Co-infection by multiple parasites is common within individuals. Interactions between co-infecting parasites include resource competition, direct competition and immune-mediated interactions and each are likely to alter the dynamics of single parasites. We posit that co-infection is a driver of variation in parasite establishment and growth, ultimately altering the production of parasite transmission stages. To test this hypothesis, three different treatment groups of laboratory mice were infected with the gastrointestinal helminth Heligmosomoides polygyrus, the respiratory bacterial pathogen Bordetella bronchiseptica lux(+) or co-infected with both parasites. To follow co-infection simultaneously, self-bioluminescent bacteria were used to quantify infection in vivo and in real-time, while helminth egg production was monitored in real-time using faecal samples. Co-infection resulted in high bacterial loads early in the infection (within the first 5 days) that could cause host mortality. Co-infection also produced helminth 'super-shedders'; individuals that chronically shed the helminth eggs in larger than average numbers. Our study shows that co-infection may be one of the underlying mechanisms for the often-observed high variance in parasite load and shedding rates, and should thus be taken into consideration for disease management and control. Further, using self-bioluminescent bacterial reporters allowed quantification of the progression of infection within the whole animal of the same individuals at a fine temporal scale (daily) and significantly reduced the number of animals used (by 85%) compared with experiments that do not use in vivo techniques. Thus, we present bioluminescent imaging as a novel, non-invasive tool offering great potential to be taken forward into other applications of infectious disease ecology.
Figures




References
-
- Ioannidis JPA, Cappelleri JC, Schmid CH, Lau J. 1996. Impact of epidemic and individual heterogeneity on the population distribution of disease progression rates: an example from patient populations in trials of human immunodeficiency virus infection. Am. J. Epidemiol. 144, 1074–108510.1093/oxfordjournals.aje.a008881 (doi:10.1093/oxfordjournals.aje.a008881) - DOI - DOI - PubMed
-
- Luong LT, Perkins SE, Grear DA, Rizzoli A, Hudson PJ. 2010. The relative importance of host characteristics and coinfection in generating variation in Heligmosomoides polygyrus fecundity. Parasitology 137, 1003–101210.1017/S0031182009991892 (doi:10.1017/S0031182009991892) - DOI - DOI - PubMed
-
- Woolhouse MEJ, et al. 1997. Heterogeneities in the transmission of infectious agents: implications for the design of control programs. Proc. Natl Acad. Sci. USA 94, 338–34210.1073/pnas.94.1.338 (doi:10.1073/pnas.94.1.338) - DOI - DOI - PMC - PubMed
-
- Perkins SE, Cattadori IM, Tagliapietra V, Rizzoli AP, Hudson PJ. 2003. Empirical key hosts in persistence of a tick-borne disease. Int. J. Parasitol. 33, 909–91710.1016/S0020-7519(03)00128-0 (doi:10.1016/S0020-7519(03)00128-0) - DOI - DOI - PubMed
-
- Chase-Topping M, Gally D, Low C, Matthews L, Woolhouse M. 2008. Super-shedding and the link between human infection and livestock carriage of Escherichia coli O157. Nat. Rev. Microbiol. 12, 904–91210.1038/nrmicro2029 (doi:10.1038/nrmicro2029) - DOI - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
