

Microtubule attachment and spindle assembly checkpoint signalling at the kinetochore
- PMID: 23258294
- PMCID: PMC3762224
- DOI: 10.1038/nrm3494
Microtubule attachment and spindle assembly checkpoint signalling at the kinetochore
Abstract
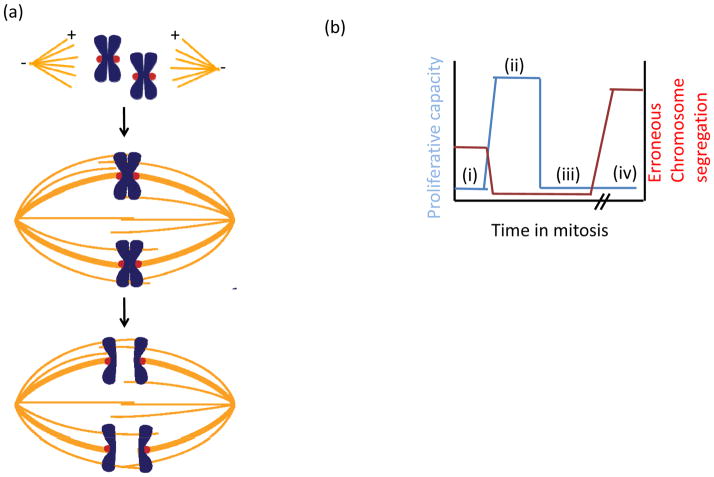
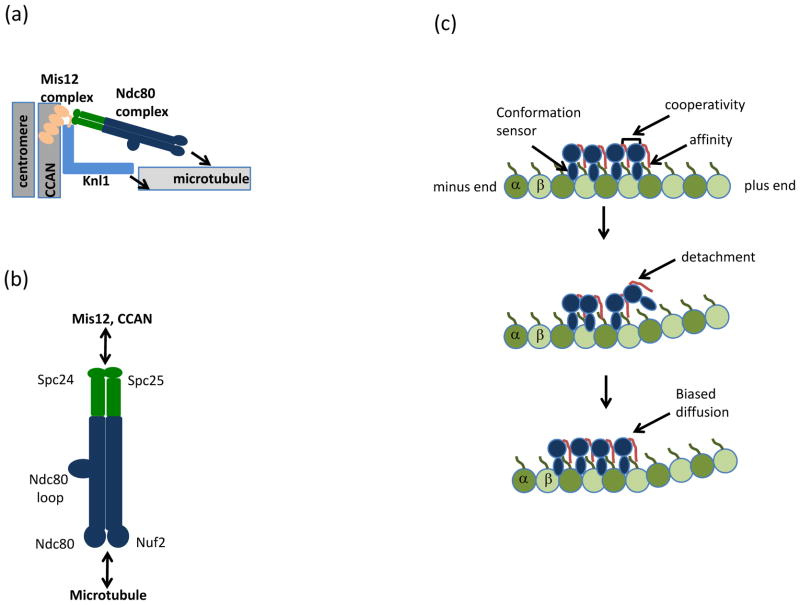
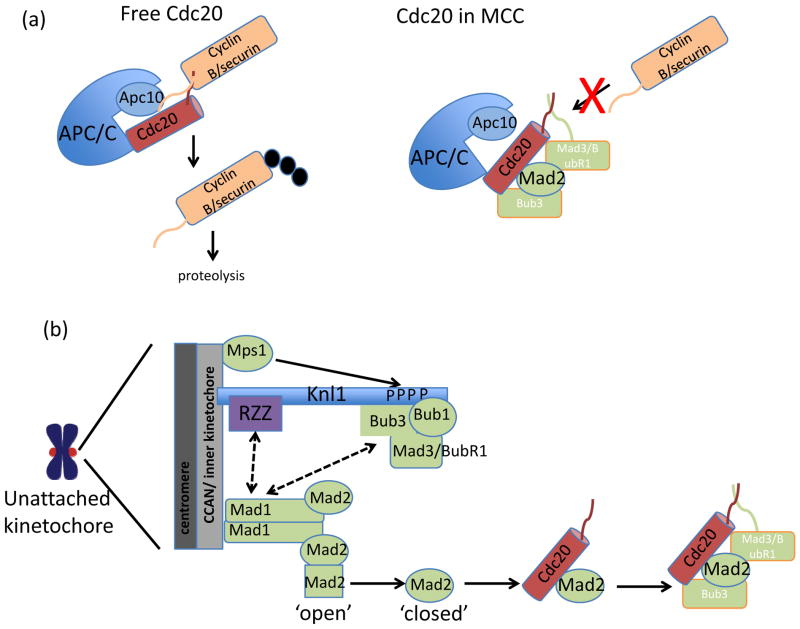
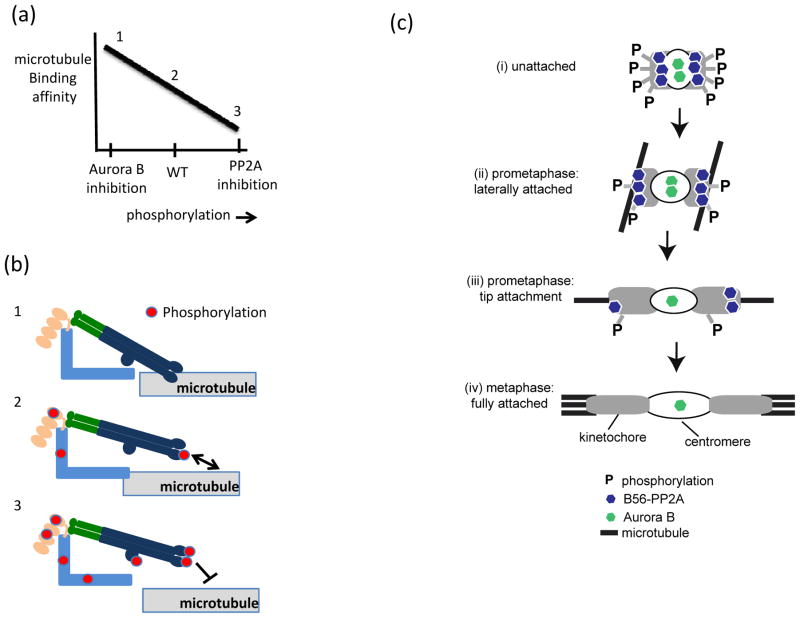
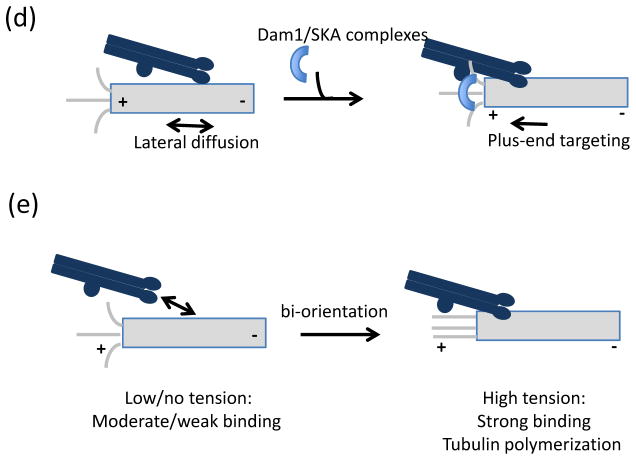
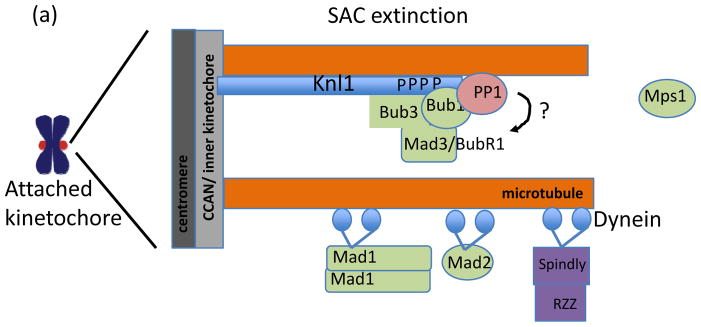
In eukaryotes, chromosome segregation during cell division is facilitated by the kinetochore, a multiprotein structure that is assembled on centromeric DNA. The kinetochore attaches chromosomes to spindle microtubules, modulates the stability of these attachments and relays the microtubule-binding status to the spindle assembly checkpoint (SAC), a cell cycle surveillance pathway that delays chromosome segregation in response to unattached kinetochores. Recent studies are shaping current thinking on how each of these kinetochore-centred processes is achieved, and how their integration ensures faithful chromosome segregation, focusing on the essential roles of kinase-phosphatase signalling and the microtubule-binding KMN protein network.
Figures

References
-
- McAinsh AD, Meraldi P. The CCAN complex: linking centromere specification to control of kinetochore-microtubule dynamics. Semin Cell Dev Biol. 2011;22:946–52. - PubMed
-
- Cheeseman IM, Chappie JS, Wilson-Kubalek EM, Desai A. The conserved KMN network constitutes the core microtubule-binding site of the kinetochore. Cell. 2006;127:983–97. A seminal paper that identifies the KMN network as a conserved essential microtubule-binding activity at the kinetochore. - PubMed
-
- Gordon DJ, Resio B, Pellman D. Causes and consequences of aneuploidy in cancer. Nat Rev Genet. 2012;13:189–203. - PubMed
-
- Lengauer C, Kinzler KW, Vogelstein B. Genetic instabilities in human cancers. Nature. 1998;396:643–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
