

Ligand-dependent conformational changes in the clamp region of the cardiac ryanodine receptor
- PMID: 23258540
- PMCID: PMC3567658
- DOI: 10.1074/jbc.M112.427864
Ligand-dependent conformational changes in the clamp region of the cardiac ryanodine receptor
Abstract
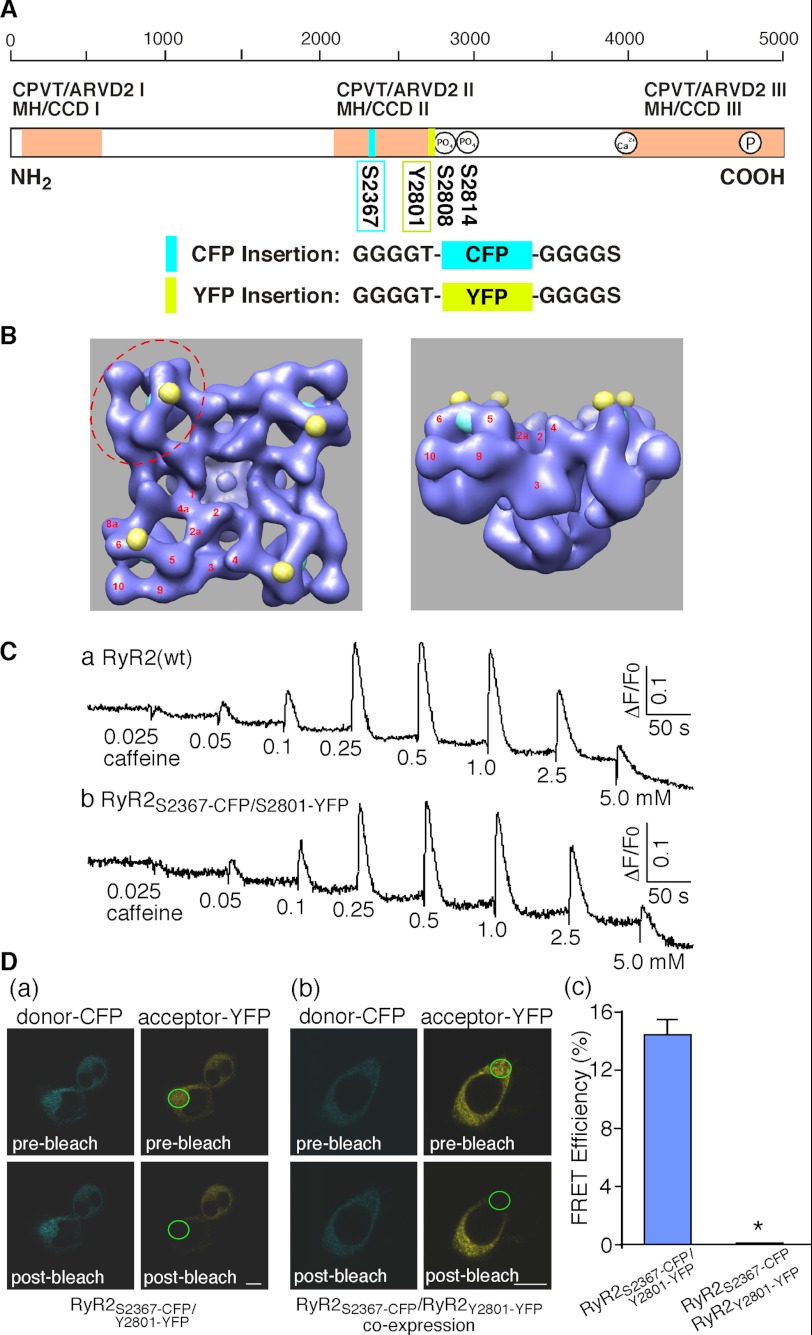
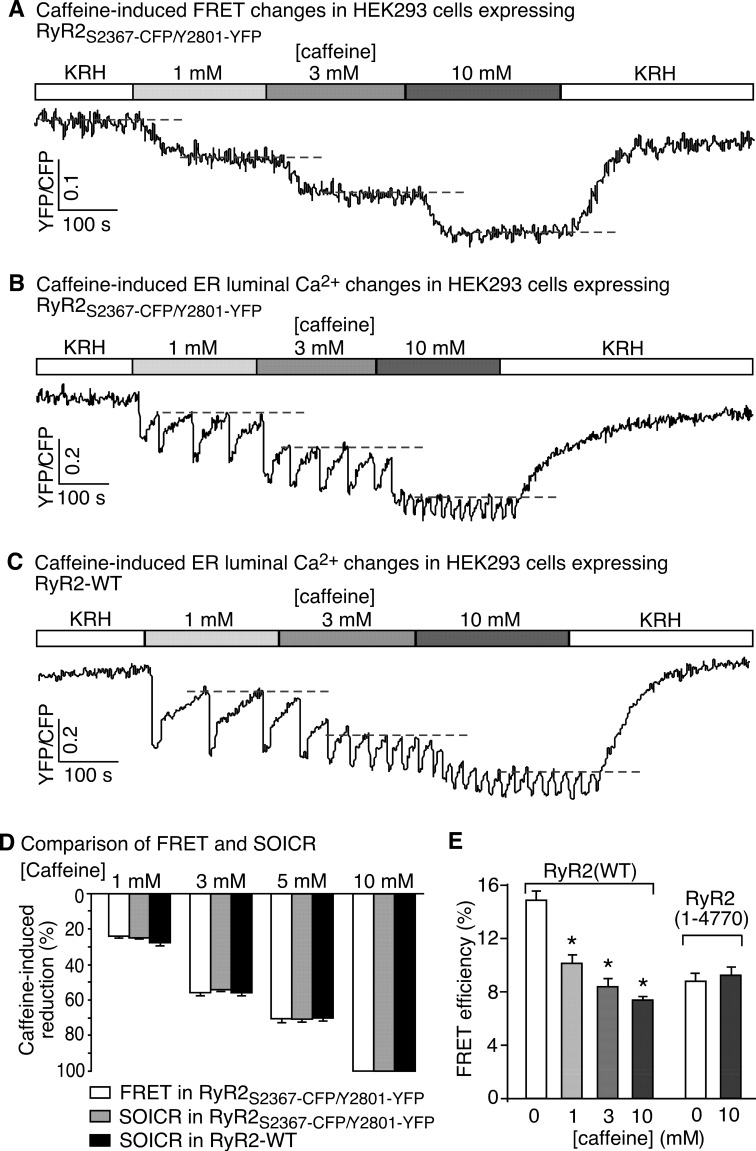
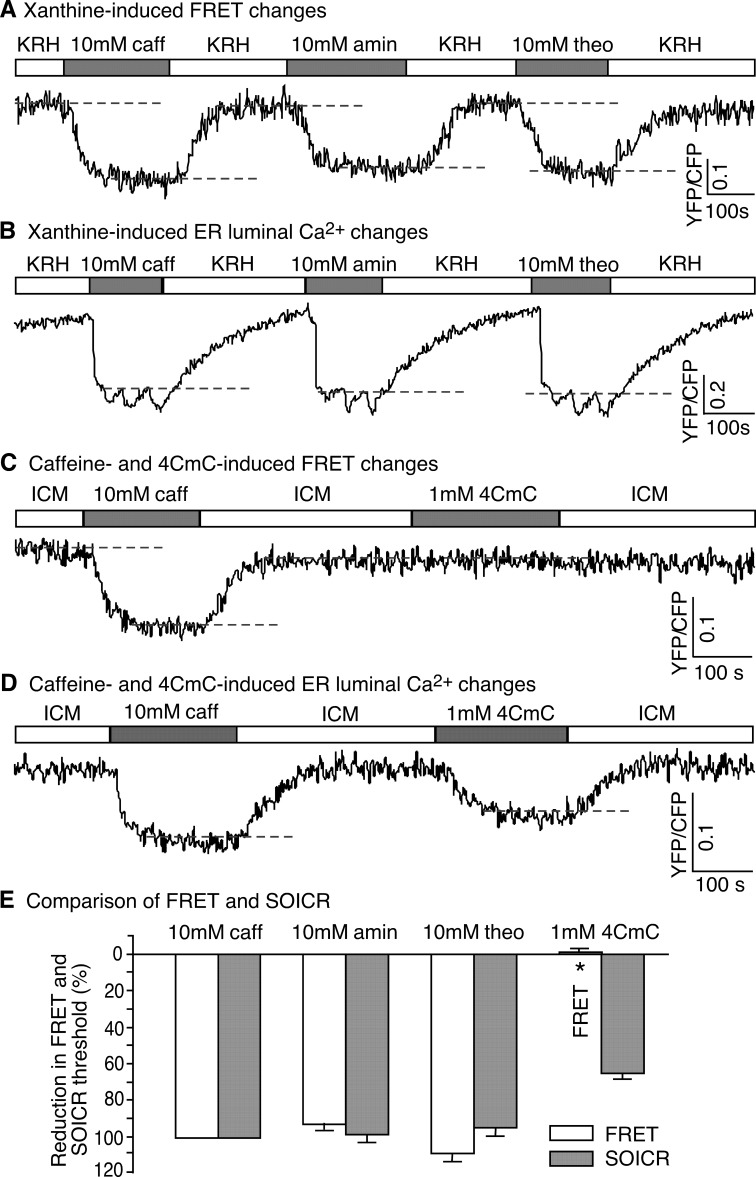
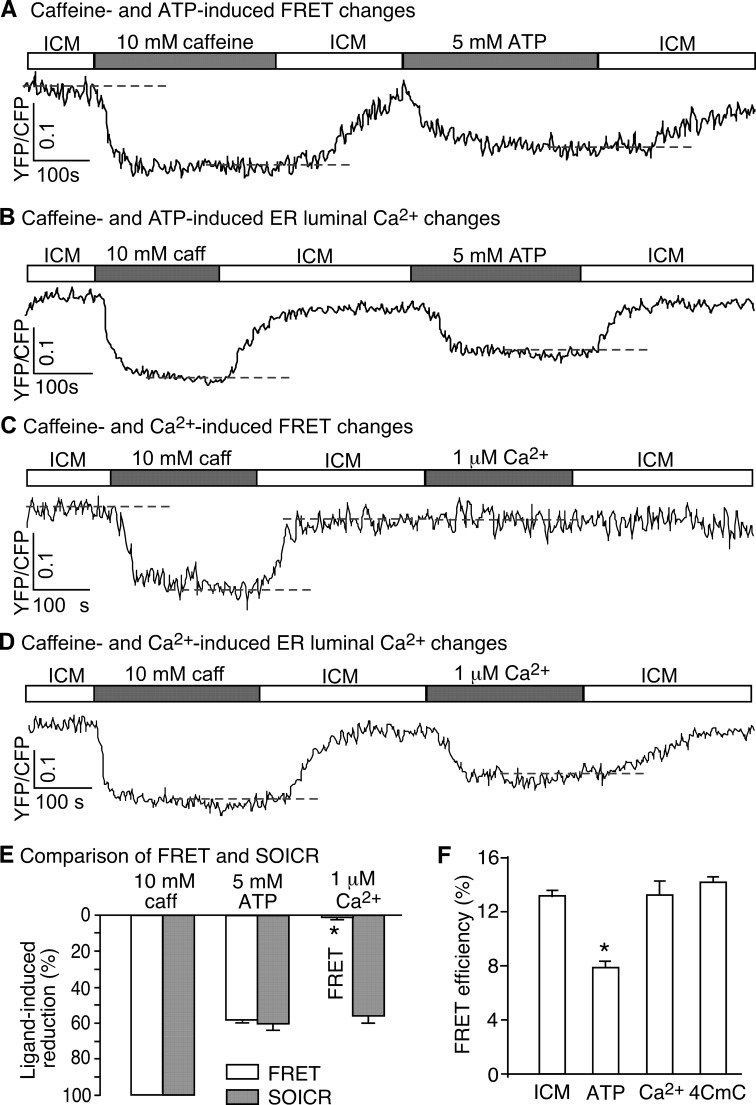
Global conformational changes in the three-dimensional structure of the Ca(2+) release channel/ryanodine receptor (RyR) occur upon ligand activation. A number of ligands are able to activate the RyR channel, but whether these structurally diverse ligands induce the same or different conformational changes in the channel is largely unknown. Here we constructed a fluorescence resonance energy transfer (FRET)-based probe by inserting a CFP after residue Ser-2367 and a YFP after residue Tyr-2801 in the cardiac RyR (RyR2) to yield a CFP- and YFP-dual labeled RyR2 (RyR2(Ser-2367-CFP/Tyr-2801-YFP)). Both of these insertion sites have previously been mapped to the "clamp" region in the four corners of the square-shaped cytoplasmic assembly of the three-dimensional structure of RyR2. Using this novel FRET probe, we monitored the extent of conformational changes in the clamp region of RyR2(Ser-2367-CFP/Tyr-2801-YFP) induced by various ligands. We also monitored the extent of Ca(2+) release induced by the same ligands in HEK293 cells expressing RyR2(Ser-2367-CFP/Tyr-2801-YFP). We detected conformational changes in the clamp region for the ligands caffeine, aminophylline, theophylline, ATP, and ryanodine but not for Ca(2+) or 4-chloro-m-cresol, although they all induced Ca(2+) release. Interestingly, caffeine is able to induce further conformational changes in the clamp region of the ryanodine-modified channel, suggesting that ryanodine does not lock RyR in a fixed conformation. Our data demonstrate that conformational changes in the clamp region of RyR are ligand-dependent and suggest the existence of multiple ligand dependent RyR activation mechanisms associated with distinct conformational changes.
Figures

References
-
- Bers D. M. (2002) Cardiac excitation-contraction coupling. Nature 415, 198–205 - PubMed
-
- Berridge M. J., Bootman M. D., Roderick H. L. (2003) Calcium signaling. Dynamics, homeostasis, and remodelling. Nat. Rev. Mol. Cell Biol. 4, 517–529 - PubMed
-
- Fill M., Copello J. A. (2002) Ryanodine receptor calcium release channels. Physiol. Rev. 82, 893–922 - PubMed
-
- Meissner G. (2004) Molecular regulation of cardiac ryanodine receptor ion channel. Cell Calcium 35, 621–628 - PubMed
-
- Zucchi R., Ronca-Testoni S. (1997) The sarcoplasmic reticulum Ca2+ channel/ryanodine receptor. Modulation by endogenous effectors, drugs, and disease states. Pharmacol. Rev. 49, 1–51 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
