

Anterograde trafficking of neurotrophin-3 in the adult olfactory system in vivo
- PMID: 23261763
- PMCID: PMC3570701
- DOI: 10.1016/j.expneurol.2012.12.010
Anterograde trafficking of neurotrophin-3 in the adult olfactory system in vivo
Abstract
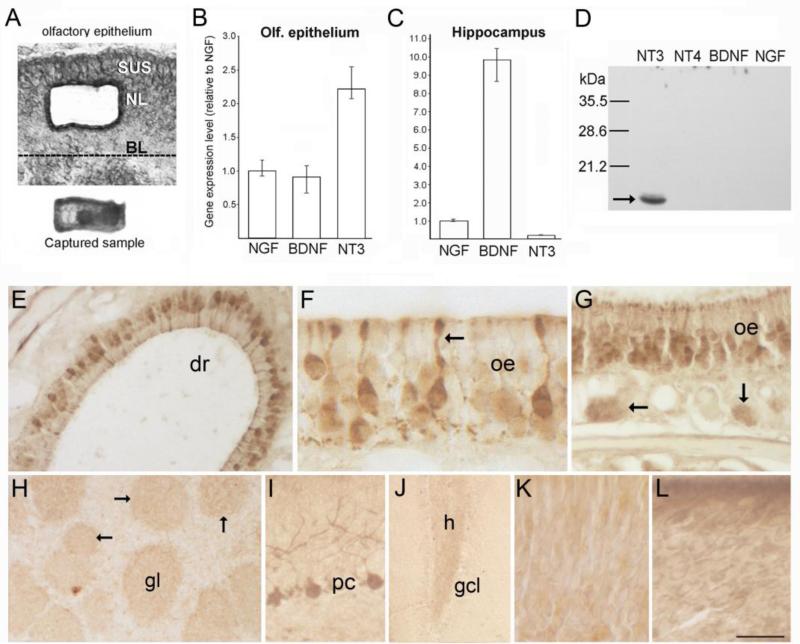
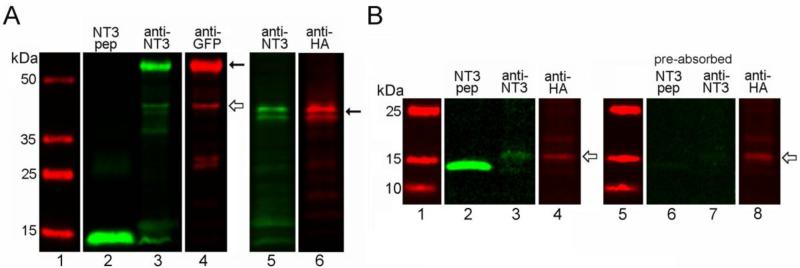
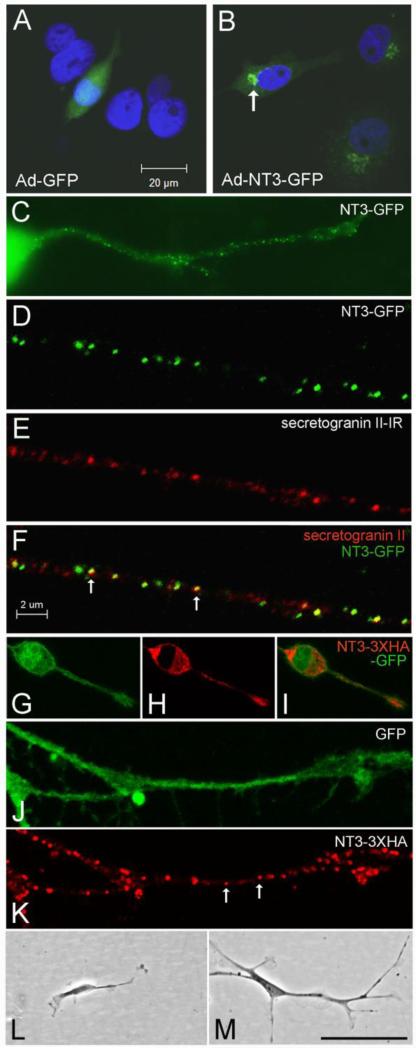
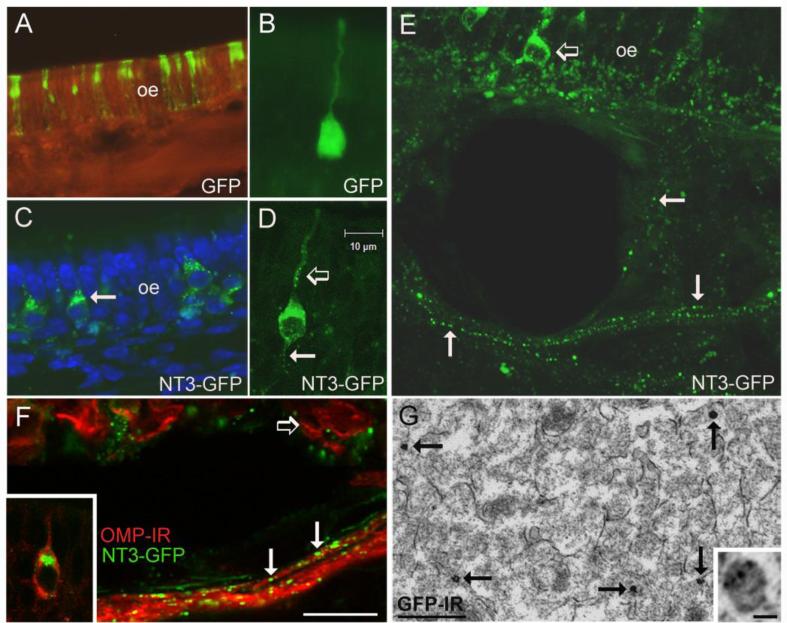
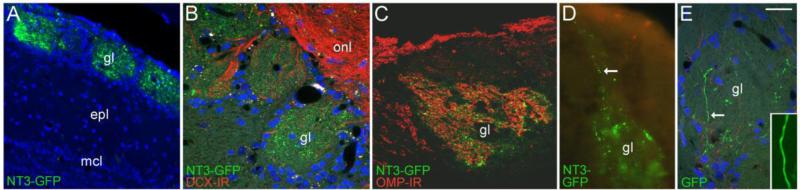
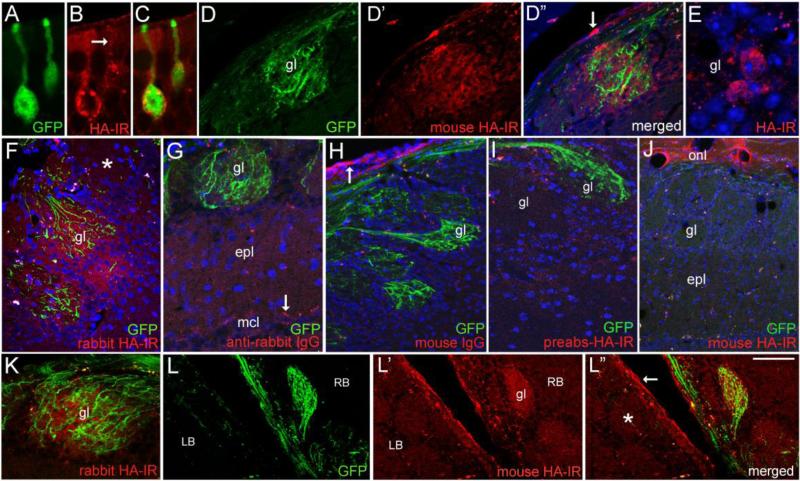
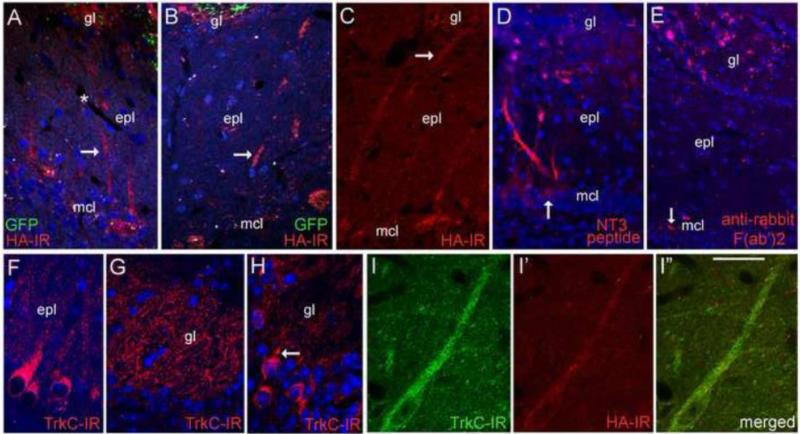
The olfactory system continuously incorporates new neurons into functional circuits throughout life. Axons from olfactory sensory neurons (OSNs) in the nasal cavity synapse on mitral, tufted and periglomerular (PG) cells in the main olfactory bulb, and low levels of turnover within the OSN population results in ingrowth of new axons under normal physiological conditions. Subpopulations of bulb interneurons are continually eliminated by apoptosis, and are replaced by new neurons derived from progenitors in the adult forebrain subventricular zone. Integration of new neurons, including PG cells that are contacted by sensory axons, leads to ongoing reorganization of adult olfactory bulb circuits. The mechanisms regulating this adaptive structural plasticity are not all known, but the process is reminiscent of early nervous system development. Neurotrophic factors have well-established roles in controlling neuronal survival and connectivity during development, leading to speculation that trophic interactions between OSNs and their target bulb neurons may mediate some of these same processes in adults. A number of different trophic factors and their cognate receptors are expressed in the adult olfactory pathway. Neurotrophin-3 (NT3) is among these, as reflected by beta-galactosidase expression in transgenic reporter mice expressing lacZ under the NT3 promoter. Using a combination of approaches, including immunocytochemistry, real-time PCR of laser-captured RNA, and adenovirus-mediated gene transfer of NT3 fusion peptides in vivo, we demonstrate that OSNs express and anterogradely transport NT3 to the olfactory bulb. We additionally observe that in mice treated with adenovirus encoding NT3 tagged with hemagglutinin (HA), a subset of bulb neurons expressing the TrkC neurotrophin receptor are immunoreactive for HA, suggesting their acquisition of the fusion peptide from infected sensory neurons. Our results therefore provide evidence that OSNs may serve as an afferent source of trophic signals for the adult mouse olfactory bulb.
Published by Elsevier Inc.
Figures
Similar articles
-
BDNF gene replacement reveals multiple mechanisms for establishing neurotrophin specificity during sensory nervous system development.Development. 2003 Apr;130(8):1479-91. doi: 10.1242/dev.00378. Development. 2003. PMID: 12620975
-
Structure and emergence of specific olfactory glomeruli in the mouse.J Neurosci. 2001 Dec 15;21(24):9713-23. doi: 10.1523/JNEUROSCI.21-24-09713.2001. J Neurosci. 2001. PMID: 11739580 Free PMC article.
-
Development and topography of the lateral olfactory tract in the mouse: imaging by genetically encoded and injected fluorescent markers.J Neurobiol. 2006 Jul;66(8):835-46. doi: 10.1002/neu.20266. J Neurobiol. 2006. PMID: 16673392
-
Classical and novel directions in neurotrophin transport and research: anterograde transport of brain-derived neurotrophic factor by sensory neurons.Microsc Res Tech. 1999 May 15-Jun 1;45(4-5):225-32. doi: 10.1002/(SICI)1097-0029(19990515/01)45:4/5<225::AID-JEMT6>3.0.CO;2-1. Microsc Res Tech. 1999. PMID: 10383115 Review.
-
Receptor guanylyl cyclases in mammalian olfactory function.Mol Cell Biochem. 2010 Jan;334(1-2):191-7. doi: 10.1007/s11010-009-0325-9. Epub 2009 Nov 26. Mol Cell Biochem. 2010. PMID: 19941039 Free PMC article. Review.
Cited by
-
Reduction of Proliferating Olfactory Cells and Low Expression of Extracellular Matrix Genes Are Hallmarks of the Aged Olfactory Mucosa.Front Aging Neurosci. 2018 Mar 27;10:86. doi: 10.3389/fnagi.2018.00086. eCollection 2018. Front Aging Neurosci. 2018. PMID: 29636678 Free PMC article.
-
Cellular Diversity and Regeneration in Taste Buds.Curr Opin Physiol. 2021 Apr;20:146-153. doi: 10.1016/j.cophys.2021.01.003. Epub 2021 Jan 12. Curr Opin Physiol. 2021. PMID: 33615087 Free PMC article.
-
Cigarette Smoke Delays Regeneration of the Olfactory Epithelium in Mice.Neurotox Res. 2016 Aug;30(2):213-24. doi: 10.1007/s12640-016-9617-5. Epub 2016 Mar 22. Neurotox Res. 2016. PMID: 27003941
-
COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2?Viruses. 2021 Nov 4;13(11):2225. doi: 10.3390/v13112225. Viruses. 2021. PMID: 34835030 Free PMC article. Review.
-
Differential localization of NT-3 and TrpM5 in glomeruli of the olfactory bulb of mice.J Comp Neurol. 2014 Jun 1;522(8):1929-40. doi: 10.1002/cne.23512. J Comp Neurol. 2014. PMID: 24288162 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
