

Interactions of Anopheles gambiae odorant-binding proteins with a human-derived repellent: implications for the mode of action of n,n-diethyl-3-methylbenzamide (DEET)
- PMID: 23261834
- PMCID: PMC3567696
- DOI: 10.1074/jbc.M112.436386
Interactions of Anopheles gambiae odorant-binding proteins with a human-derived repellent: implications for the mode of action of n,n-diethyl-3-methylbenzamide (DEET)
Abstract
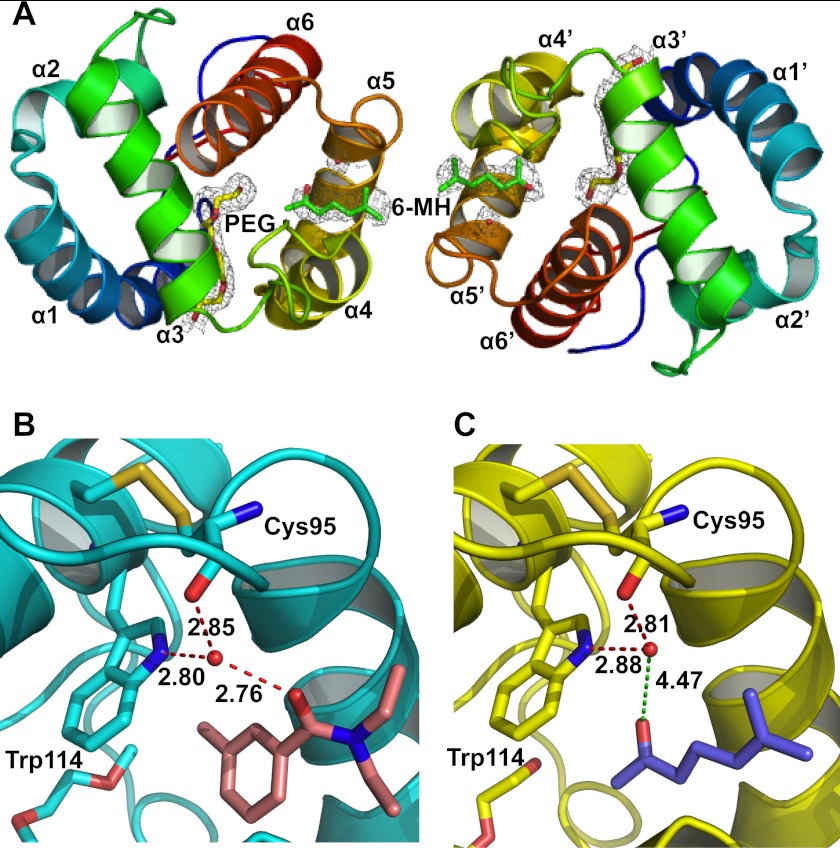
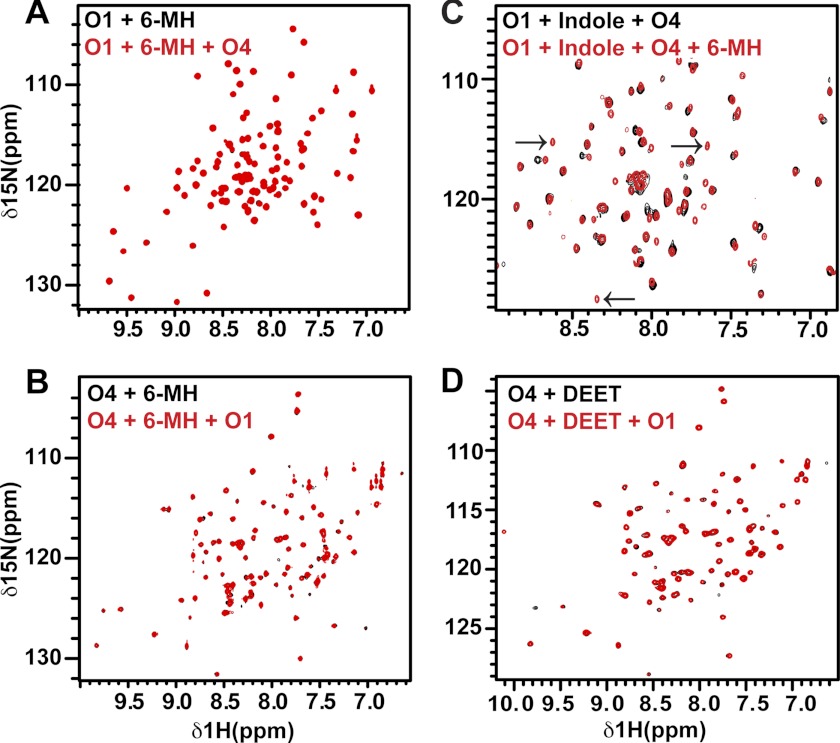
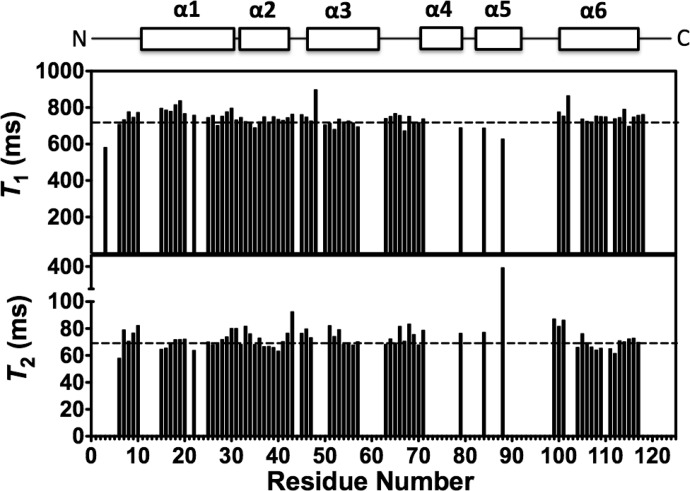
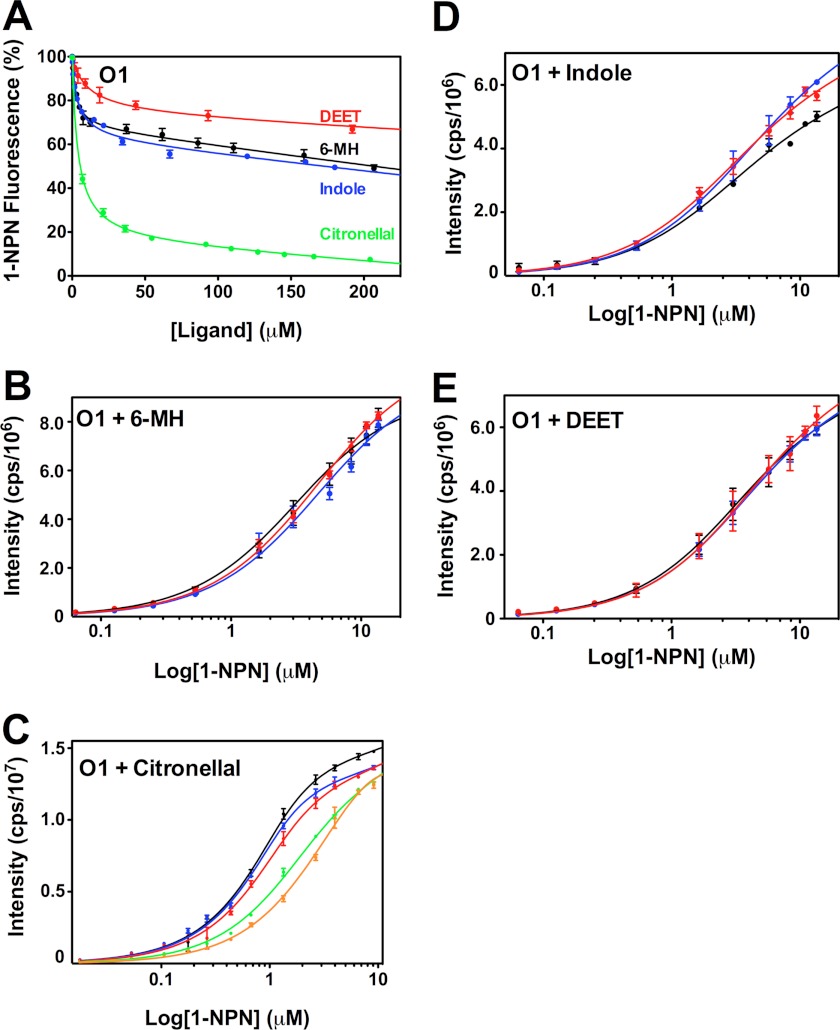
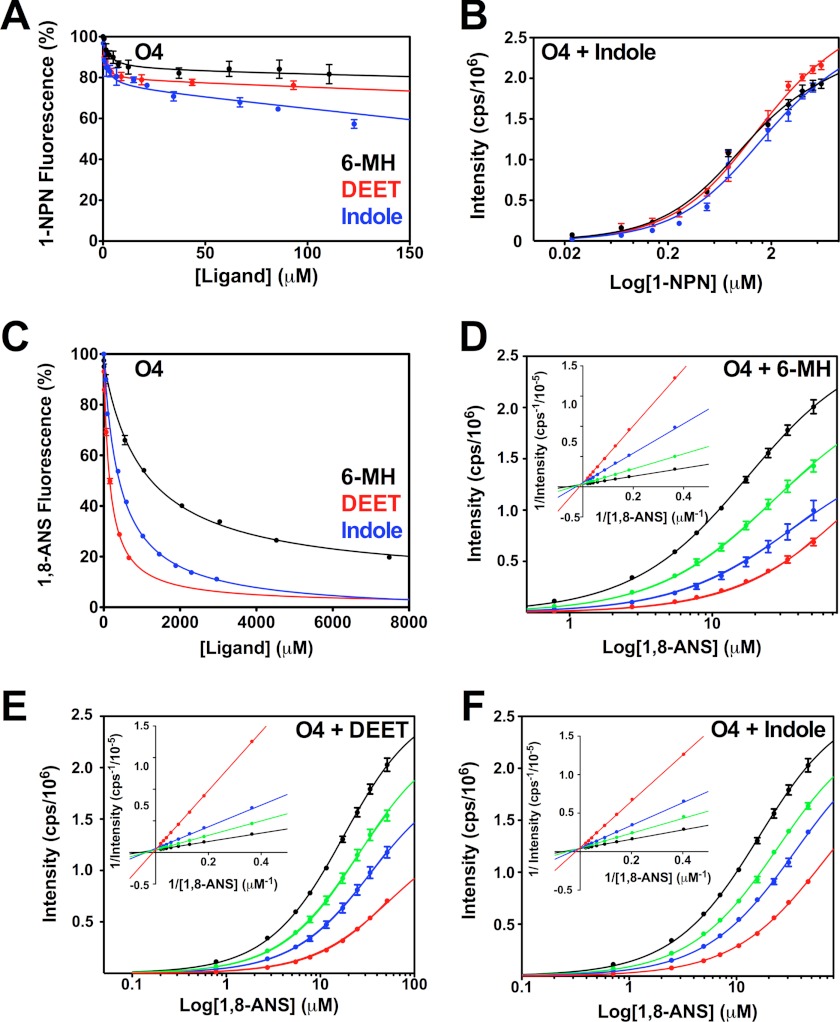
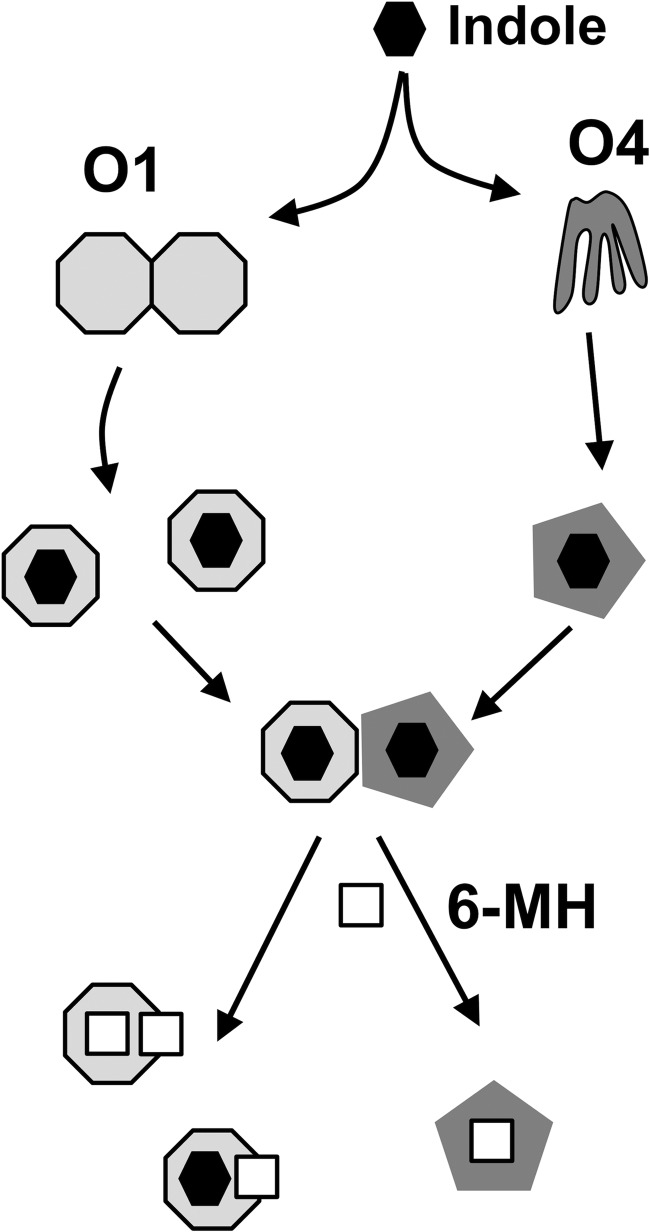
The Anopheles gambiae mosquito, which is the vector for Plasmodium falciparum malaria, uses a series of olfactory cues emanating from human sweat to select humans as their source for a blood meal. Perception of these odors within the mosquito olfactory system involves the interplay of odorant-binding proteins (OBPs) and odorant receptors and disrupting the normal responses to those odorants that guide mosquito-human interactions represents an attractive approach to prevent the transmission of malaria. Previously, it has been shown that DEET targets multiple components of the olfactory system, including OBPs and odorant receptors. Here, we present the crystal structure of A. gambiae OBP1 (OBP1) in the complex it forms with a natural repellent 6-methyl-5-heptene-2-one (6-MH). We find that 6-MH binds to OBP1 at exactly the same site as DEET. However, key interactions with a highly conserved water molecule that are proposed to be important for DEET binding are not involved in binding of 6-MH. We show that 6-MH and DEET can compete for the binding of attractive odorants and in doing so disrupt the interaction that OBP1 makes with OBP4. We further show that 6-MH and DEET can bind simultaneously to OBPs with other ligands. These results suggest that the successful discovery of novel reagents targeting OBP function requires knowledge about the specific mechanism of binding to the OBP rather than their binding affinity.
Figures
References
-
- Dondorp A. M., Nosten F., Yi P., Das D., Phyo A. P., Tarning J., Lwin K. M., Ariey F., Hanpithakpong W., Lee S. J., Ringwald P., Silamut K., Imwong M., Chotivanich K., Lim P., Herdman T., An S. S., Yeung S., Singhasivanon P., Day N. P., Lindegardh N., Socheat D., White N. J. (2009) Artemisinin resistance in Plasmodium falciparum malaria. New Engl. J. Med. 361, 455–467 - PMC - PubMed
-
- Trape J. F., Rogier C. (1996) Combating malaria morbidity and mortality by reducing transmission. Parasitol. Today 12, 236–240 - PubMed
-
- Hallem E. A., Nicole Fox A., Zwiebel L. J., Carlson J. R. (2004) Olfaction: mosquito receptor for human-sweat odorant. Nature 427, 212–213 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
