

Hierarchical accumulation of RyR post-translational modifications drives disease progression in dystrophic cardiomyopathy
- PMID: 23263329
- PMCID: PMC3583259
- DOI: 10.1093/cvr/cvs425
Hierarchical accumulation of RyR post-translational modifications drives disease progression in dystrophic cardiomyopathy
Abstract
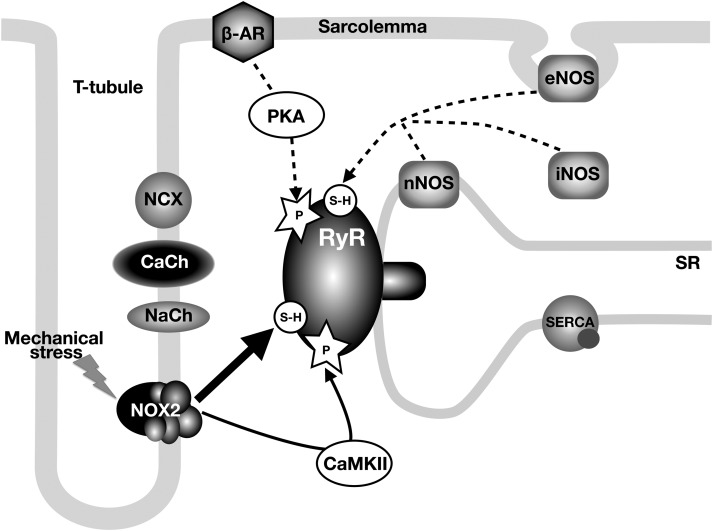
Aims: Duchenne muscular dystrophy (DMD) is a muscle disease with serious cardiac complications. Changes in Ca(2+) homeostasis and oxidative stress were recently associated with cardiac deterioration, but the cellular pathophysiological mechanisms remain elusive. We investigated whether the activity of ryanodine receptor (RyR) Ca(2+) release channels is affected, whether changes in function are cause or consequence and which post-translational modifications drive disease progression.
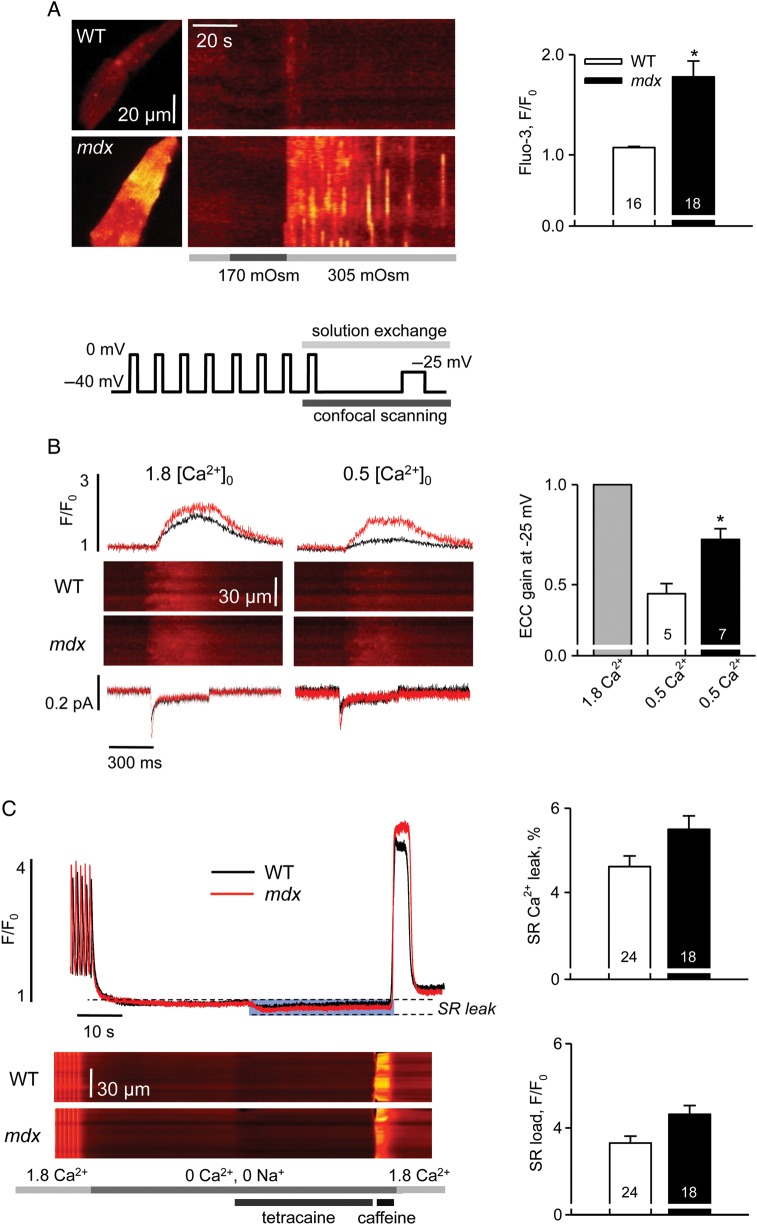
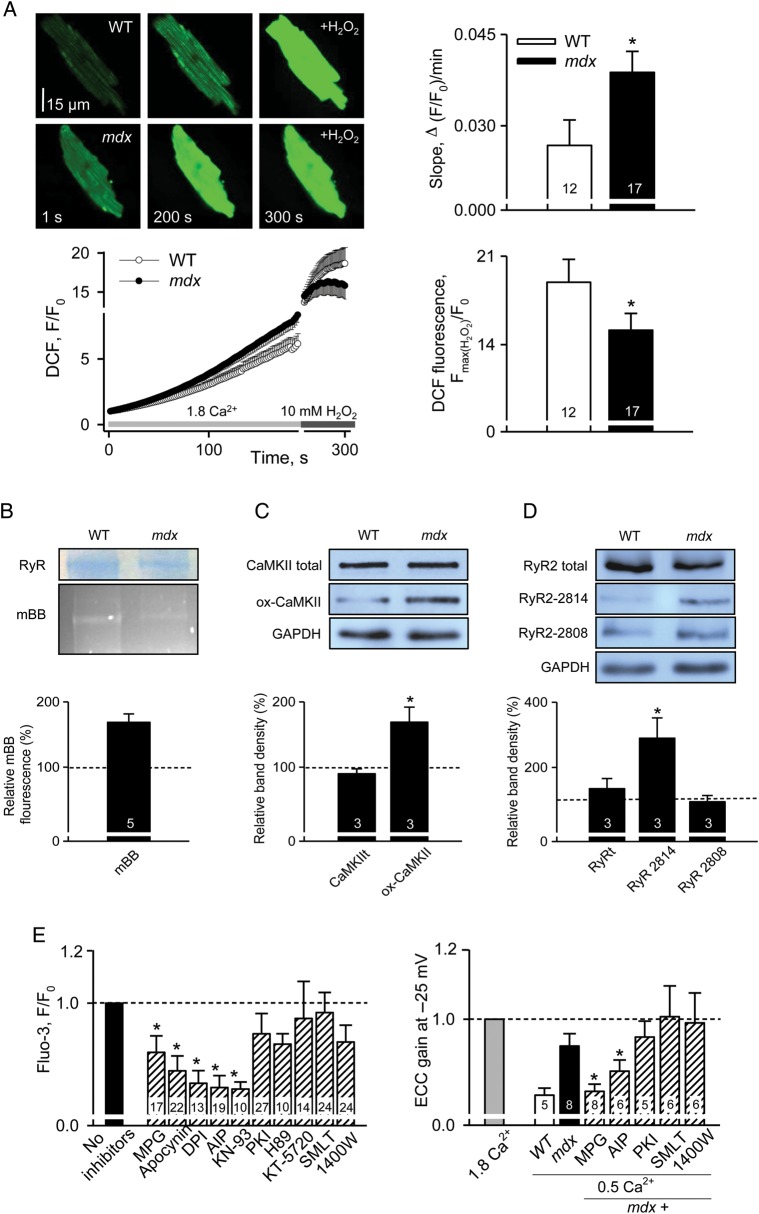
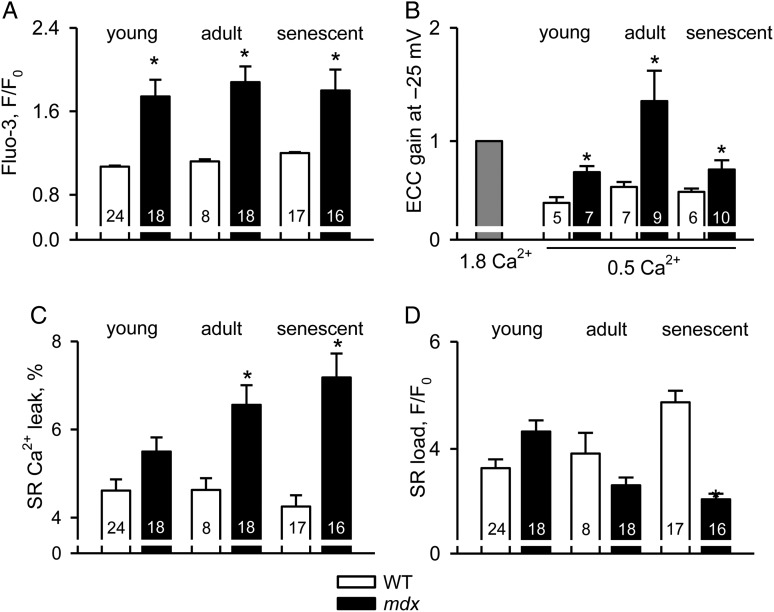
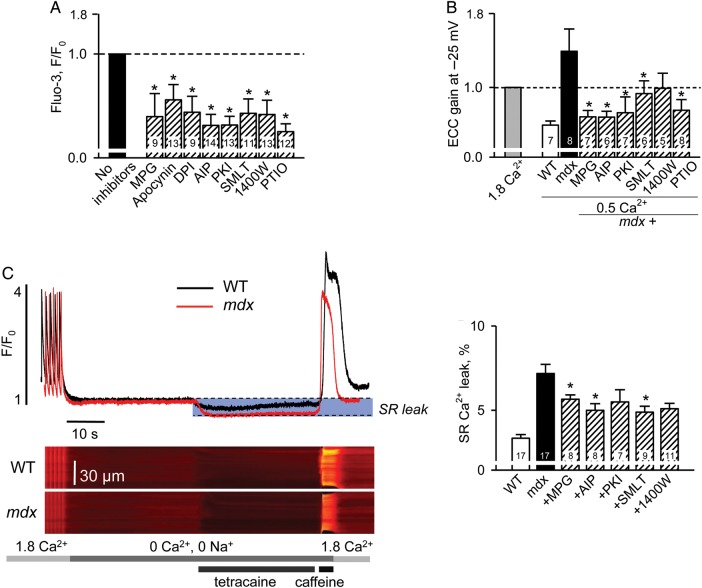
Methods and results: Electrophysiological, imaging, and biochemical techniques were used to study RyRs in cardiomyocytes from mdx mice, an animal model of DMD. Young mdx mice show no changes in cardiac performance, but do so after ∼8 months. Nevertheless, myocytes from mdx pups exhibited exaggerated Ca(2+) responses to mechanical stress and 'hypersensitive' excitation-contraction coupling, hallmarks of increased RyR Ca(2+) sensitivity. Both were normalized by antioxidants, inhibitors of NAD(P)H oxidase and CaMKII, but not by NO synthases and PKA antagonists. Sarcoplasmic reticulum Ca(2+) load and leak were unchanged in young mdx mice. However, by the age of 4-5 months and in senescence, leak was increased and load was reduced, indicating disease progression. By this age, all pharmacological interventions listed above normalized Ca(2+) signals and corrected changes in ECC, Ca(2+) load, and leak.
Conclusion: Our findings suggest that increased RyR Ca(2+) sensitivity precedes and presumably drives the progression of dystrophic cardiomyopathy, with oxidative stress initiating its development. RyR oxidation followed by phosphorylation, first by CaMKII and later by PKA, synergistically contributes to cardiac deterioration.
Figures
References
-
- Hein S, Kostin S, Heling A, Maeno Y, Schaper J. The role of the cytoskeleton in heart failure. Cardiovasc Res. 2000;45:273–278. - PubMed
-
- Durbeej M, Campbell KP. Muscular dystrophies involving the dystrophin–glycoprotein complex: an overview of current mouse models. Curr Opin Genet Dev. 2002;12:349–361. - PubMed
-
- Finsterer J, Stöllberger C. The heart in human dystrophinopathies. Cardiology. 2003;99:1–19. - PubMed
-
- Quinlan JG, Hahn HS, Wong BL, Lorenz JN, Wenisch AS, Levin LS. Evolution of the mdx mouse cardiomyopathy: physiological and morphological findings. Neuromuscul Disord. 2004;14:491–496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
