

Both CpG methylation and activation-induced deaminase are required for the fragility of the human bcl-2 major breakpoint region: implications for the timing of the breaks in the t(14;18) translocation
- PMID: 23263985
- PMCID: PMC3623081
- DOI: 10.1128/MCB.01436-12
Both CpG methylation and activation-induced deaminase are required for the fragility of the human bcl-2 major breakpoint region: implications for the timing of the breaks in the t(14;18) translocation
Abstract
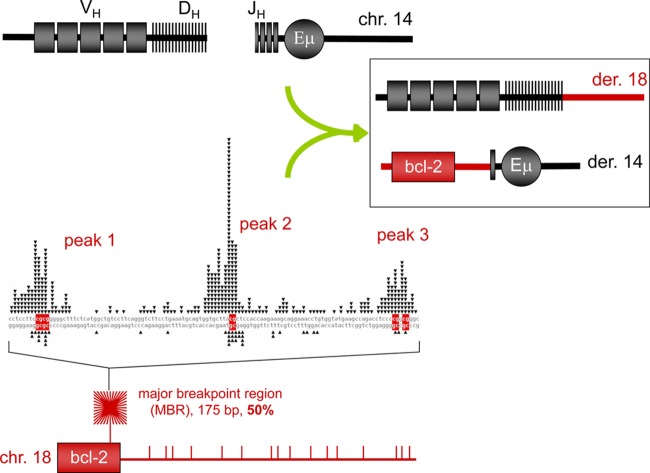
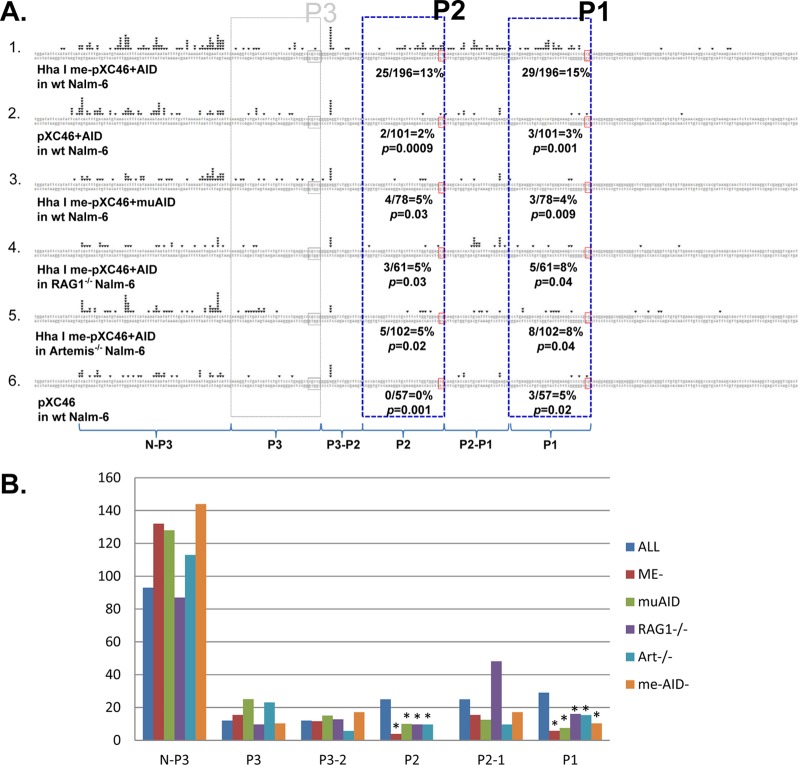
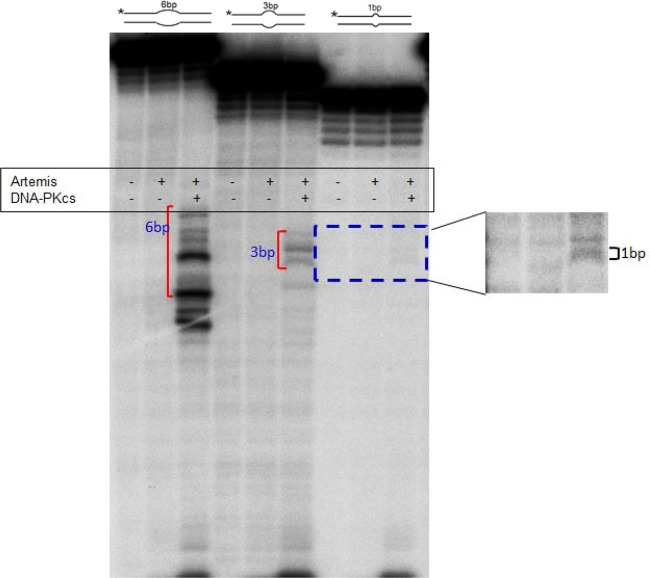
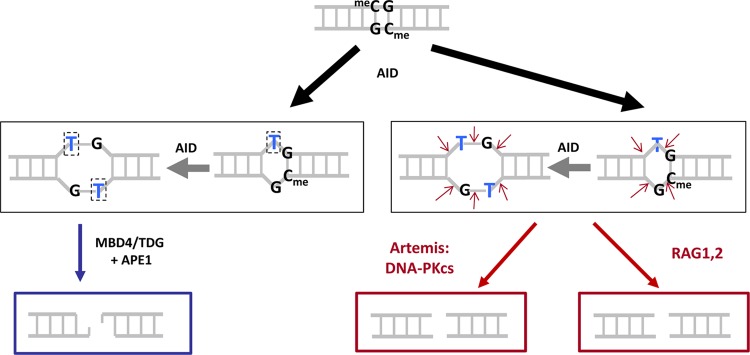
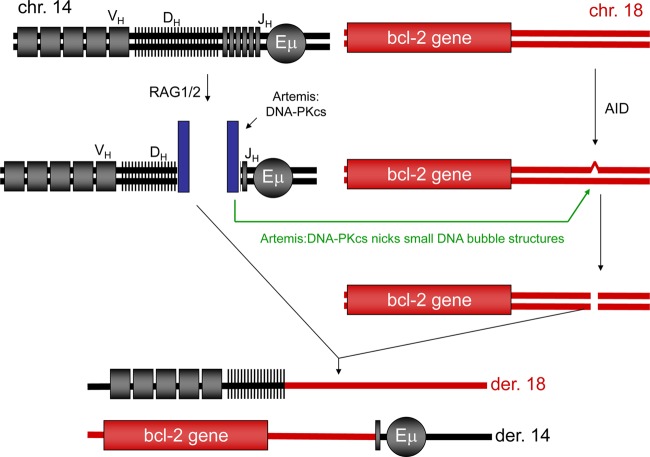
The t(14;18) chromosomal translocation typically involves breakage at the bcl-2 major breakpoint region (MBR) to cause human follicular lymphoma. A theory to explain the striking propensity of the MBR breaks at three CpG clusters within the 175-bp MBR region invoked activation-induced deaminase (AID). In a test of that theory, we used here minichromosomal substrates in human pre-B cell lines. Consistent with the essential elements of the theory, we found that the MBR breakage process is indeed highly dependent on DNA methylation at the CpG sites and highly dependent on the AID enzyme to create lesions at peak locations within the MBR. Interestingly, breakage of the phosphodiester bonds at the AID-initiated MBR lesions is RAG dependent, but, unexpectedly, most are also dependent on Artemis. We found that Artemis is capable of nicking small heteroduplex structures and is even able to nick single-base mismatches. This raises the possibility that activated Artemis, derived from the unjoined D to J(H) DNA ends at the IgH locus on chromosome 14, nicks AID-generated TG mismatches at methyl CpG sites, and this would explain why the breaks at the chromosome 18 MBR occur within the same time window as those on chromosome 14.
Figures

References
-
- Boboila C, Alt FW, Schwer B. 2012. Classical and alternative end-joining pathways for repair of lymphocyte-specific and general DNA double-strand breaks. Adv. Immunol. 116: 1–49 - PubMed
-
- Gostissa M, Alt FW, Chiarle R. 2011. Mechanisms that promote and suppress chromosomal translocations in lymphocytes. Annu. Rev. Immunol. 29: 319–350 - PubMed
-
- Hakim O, Resch W, Yamane A, Klein I, Kieffer-Kwon KR, Jankovic M, Oliveira T, Bothmer A, Voss TC, Ansarah-Sobrinho C, Mathe E, Liang G, Cobell J, Nakahashi H, Robbiani DF, Nussenzweig A, Hager GL, Nussenzweig MC, Casellas R. 2012. DNA damage defines sites of recurrent chromosomal translocations in B lymphocytes. Nature 484: 69–74 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous