

Multiple phytohormone signalling pathways modulate susceptibility of tomato plants to Alternaria alternata f. sp. lycopersici
- PMID: 23264518
- PMCID: PMC3542053
- DOI: 10.1093/jxb/ers360
Multiple phytohormone signalling pathways modulate susceptibility of tomato plants to Alternaria alternata f. sp. lycopersici
Abstract
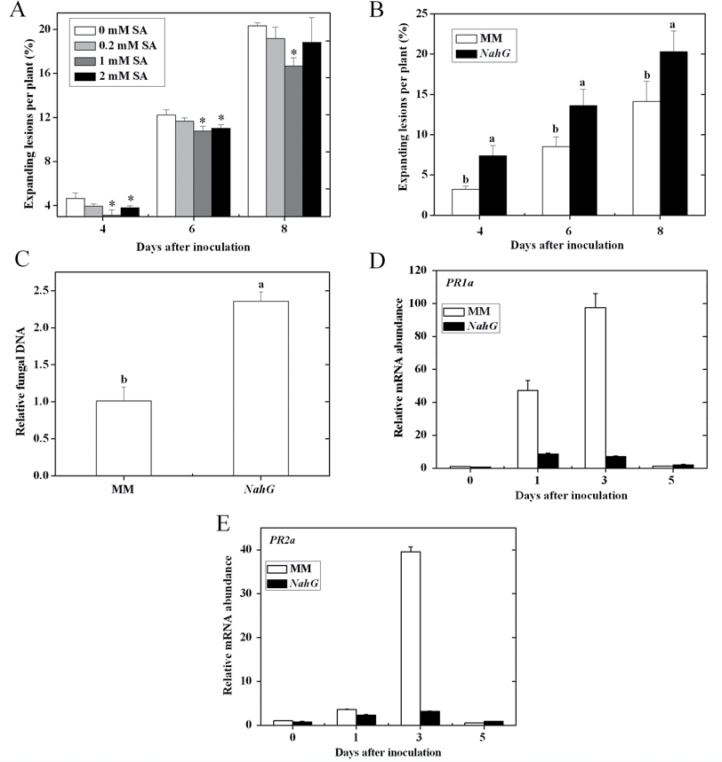
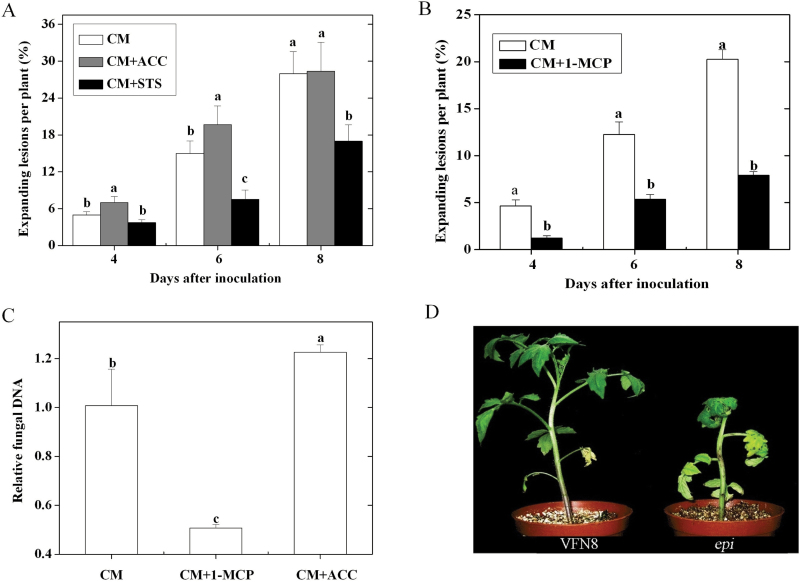
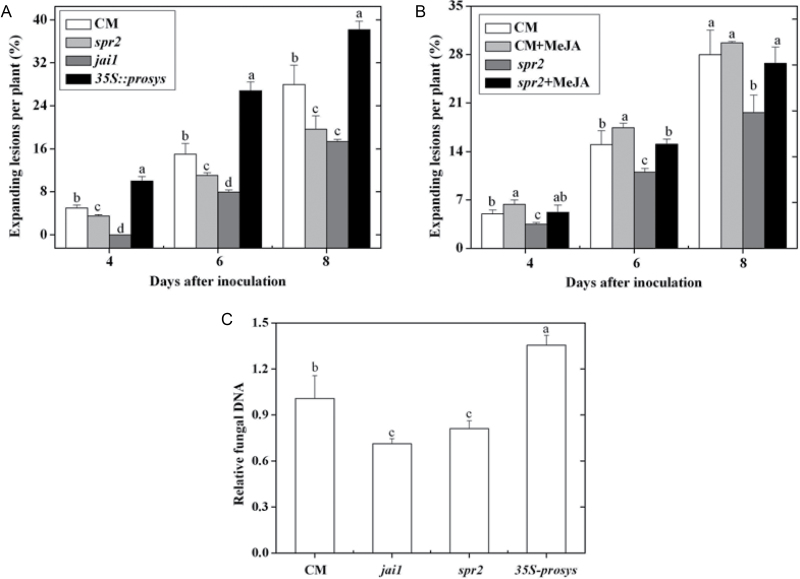
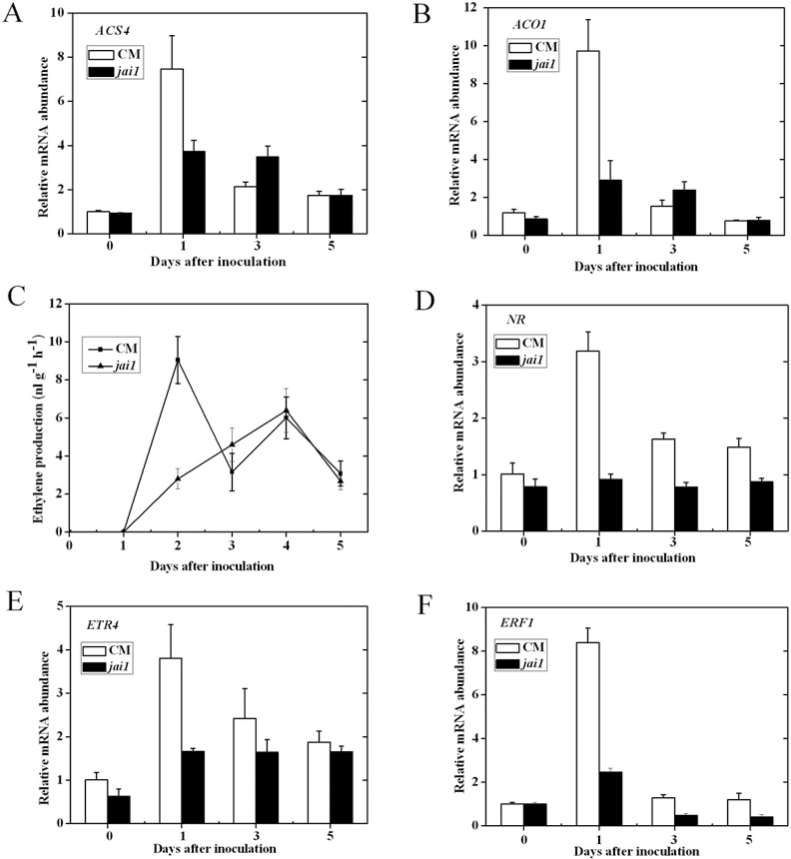
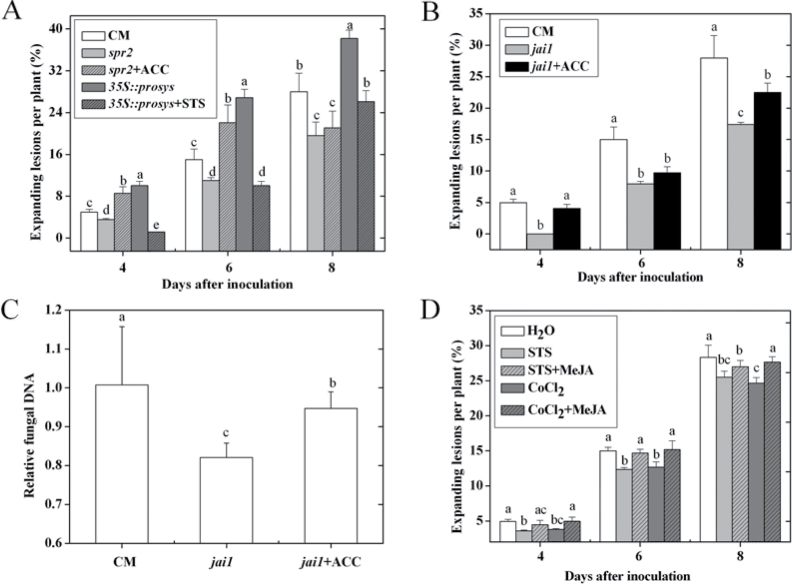
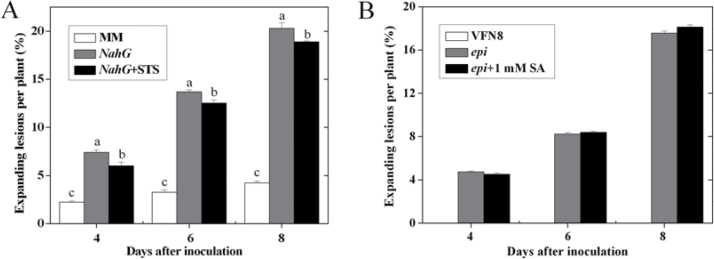
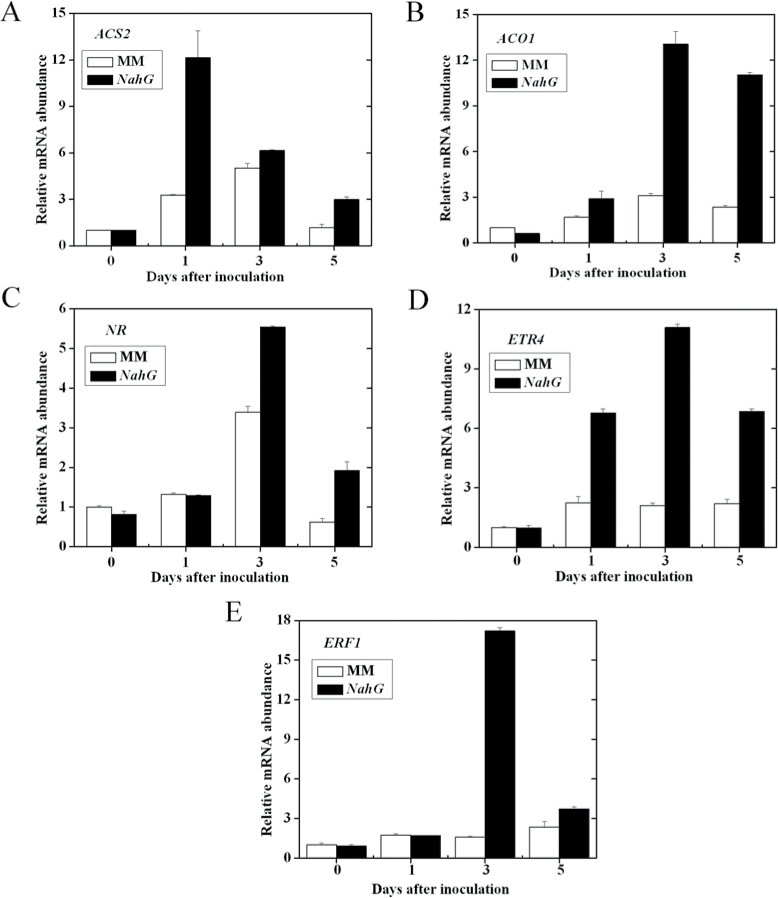
Three phytohormone molecules - ethylene (ET), jasmonic acid (JA) and salicylic acid (SA) - play key roles in mediating disease response to necrotrophic fungal pathogens. This study investigated the roles of the ET, JA, and SA pathways as well as their crosstalk during the interaction between tomato (Solanum lycopersicum) plants and a necrotrophic fungal pathogen Alternaria alternata f. sp. lycopersici (AAL). Both the ET and JASMONIC ACID INSENSITIVE1 (JAI1) receptor-dependent JA signalling pathways are necessary for susceptibility, while SA response promotes resistance to AAL infection. In addition, the role of JA in susceptibility to AAL is partly dependent on ET biosynthesis and perception, while the SA pathway enhances resistance to AAL and antagonizes the ET response. Based on these results, it is proposed that ET, JA, and SA each on their own can influence the susceptibility of tomato to AAL. Furthermore, the functions of JA and SA in susceptibility to the pathogen are correlated with the enhanced or decreased action of ET, respectively. This study has revealed the functional relationship among the three key hormone pathways in tomato defence against AAL.
Figures

Similar articles
-
Involvement of salicylic acid, ethylene and jasmonic acid signalling pathways in the susceptibility of tomato to Fusarium oxysporum.Mol Plant Pathol. 2017 Sep;18(7):1024-1035. doi: 10.1111/mpp.12559. Epub 2017 May 23. Mol Plant Pathol. 2017. PMID: 28390170 Free PMC article.
-
The involvement of jasmonates and ethylene in Alternaria alternata f. sp. lycopersici toxin-induced tomato cell death.J Exp Bot. 2011 Nov;62(15):5405-18. doi: 10.1093/jxb/err217. Epub 2011 Aug 23. J Exp Bot. 2011. PMID: 21865178 Free PMC article.
-
Critical Role of COI1-Dependent Jasmonate Pathway in AAL toxin induced PCD in Tomato Revealed by Comparative Proteomics.Sci Rep. 2016 Jun 21;6:28451. doi: 10.1038/srep28451. Sci Rep. 2016. PMID: 27324416 Free PMC article.
-
Crosstalk among Jasmonate, Salicylate and Ethylene Signaling Pathways in Plant Disease and Immune Responses.Curr Protein Pept Sci. 2015;16(5):450-61. doi: 10.2174/1389203716666150330141638. Curr Protein Pept Sci. 2015. PMID: 25824390 Review.
-
CRISPR/Cas9-Mediated Immunity in Plants Against Pathogens.Curr Issues Mol Biol. 2018;26:55-64. doi: 10.21775/cimb.026.055. Epub 2017 Sep 7. Curr Issues Mol Biol. 2018. PMID: 28879856 Review.
Cited by
-
Uncovering the mechanisms of salicylic acid-mediated abiotic stress tolerance in horticultural crops.Front Plant Sci. 2023 Aug 28;14:1226041. doi: 10.3389/fpls.2023.1226041. eCollection 2023. Front Plant Sci. 2023. PMID: 37701800 Free PMC article. Review.
-
Transcriptome analysis of the compatible interaction of tomato with Verticillium dahliae using RNA-sequencing.Front Plant Sci. 2015 Jun 8;6:428. doi: 10.3389/fpls.2015.00428. eCollection 2015. Front Plant Sci. 2015. PMID: 26106404 Free PMC article.
-
Exogenous Bio-Based 2,3-Butanediols Enhanced Abiotic Stress Tolerance of Tomato and Turfgrass under Drought or Chilling Stress.J Microbiol Biotechnol. 2022 May 28;32(5):582-593. doi: 10.4014/jmb.2201.01025. J Microbiol Biotechnol. 2022. PMID: 35484968 Free PMC article.
-
RNA-Seq derived identification of differential transcription in the chrysanthemum leaf following inoculation with Alternaria tenuissima.BMC Genomics. 2014 Jan 4;15:9. doi: 10.1186/1471-2164-15-9. BMC Genomics. 2014. PMID: 24387266 Free PMC article.
-
Does Constitutive Expression of Defense-Related Genes and Salicylic Acid Concentrations Correlate with Field Resistance of Potato to Black Scurf Disease?Bioengineering (Basel). 2023 Oct 24;10(11):1244. doi: 10.3390/bioengineering10111244. Bioengineering (Basel). 2023. PMID: 38002368 Free PMC article.
References
-
- Berrocal-Lobo M, Molina A. 2004. Ethylene response factor 1 mediates Arabidopsis resistance to the soilborne fungus Fusarium oxysporum . Molecular Plant–Microbe Interactions 17, 763–770 - PubMed
-
- Brading PA, Hammond-Kosack KE, Parr A, Jones JD. 2000. Salicylic acid is not required for Cf-2- and Cf-9-dependent resistance of tomato to Cladosporium fulvum . The Plant Journal 23, 305–318 - PubMed
-
- Brandwagt BF, Kneppers TJA, Nijkamp HJJ, Hille J. 2002. Overexpression of the tomato Asc-1 gene mediates high insensitivity to AAL toxins and fumonisin B1 in tomato hairy roots and confers resistance to Alternaria alternata f. sp. lycopersici in Nicotiana umbratica plants. Molecular Plant–Microbe Interactions 15, 35–42 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
