

Focal adhesion kinase regulates the localization and retention of pro-B cells in bone marrow microenvironments
- PMID: 23264658
- PMCID: PMC3552136
- DOI: 10.4049/jimmunol.1202639
Focal adhesion kinase regulates the localization and retention of pro-B cells in bone marrow microenvironments
Abstract
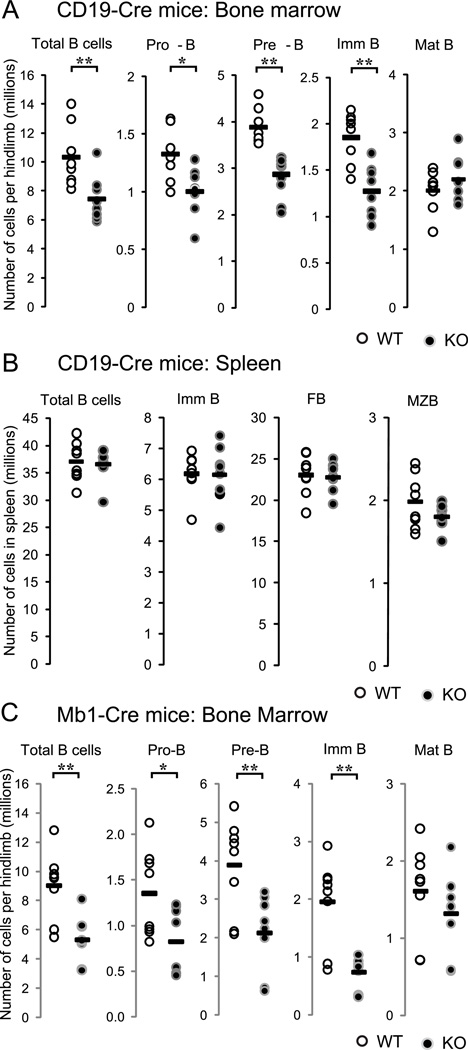
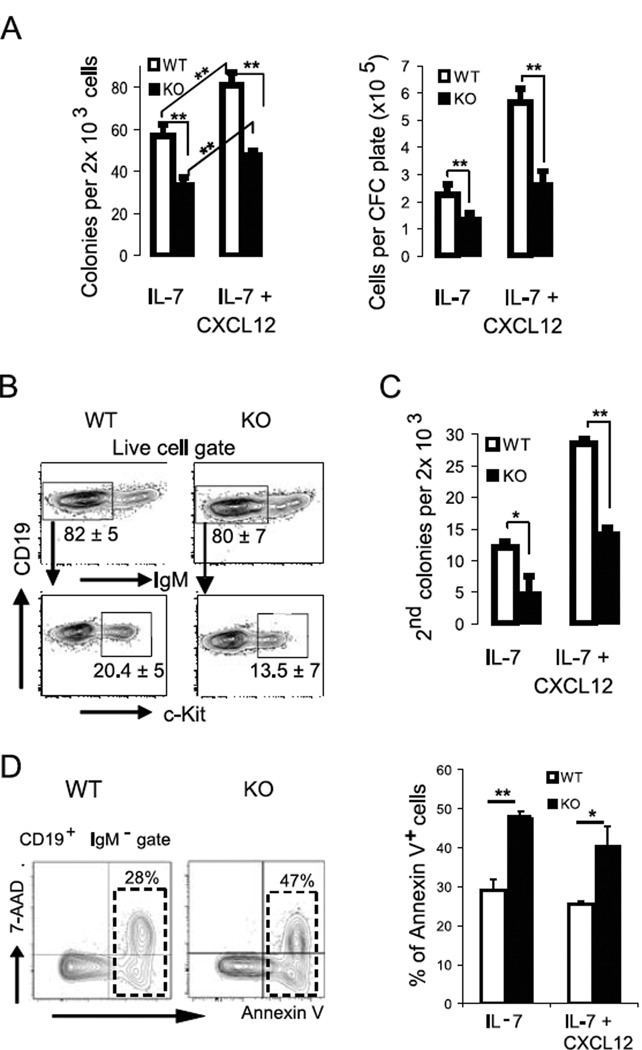
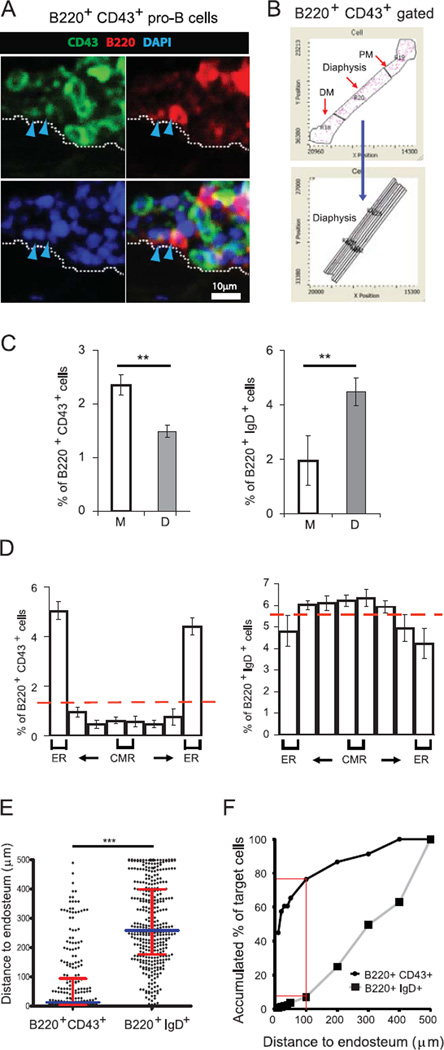
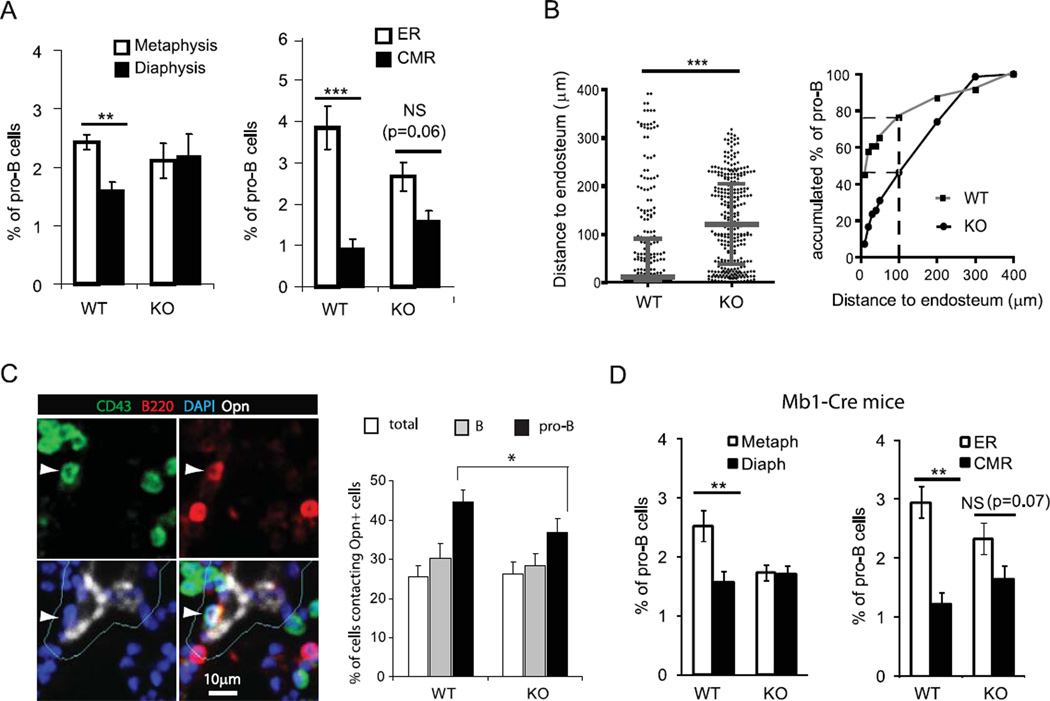
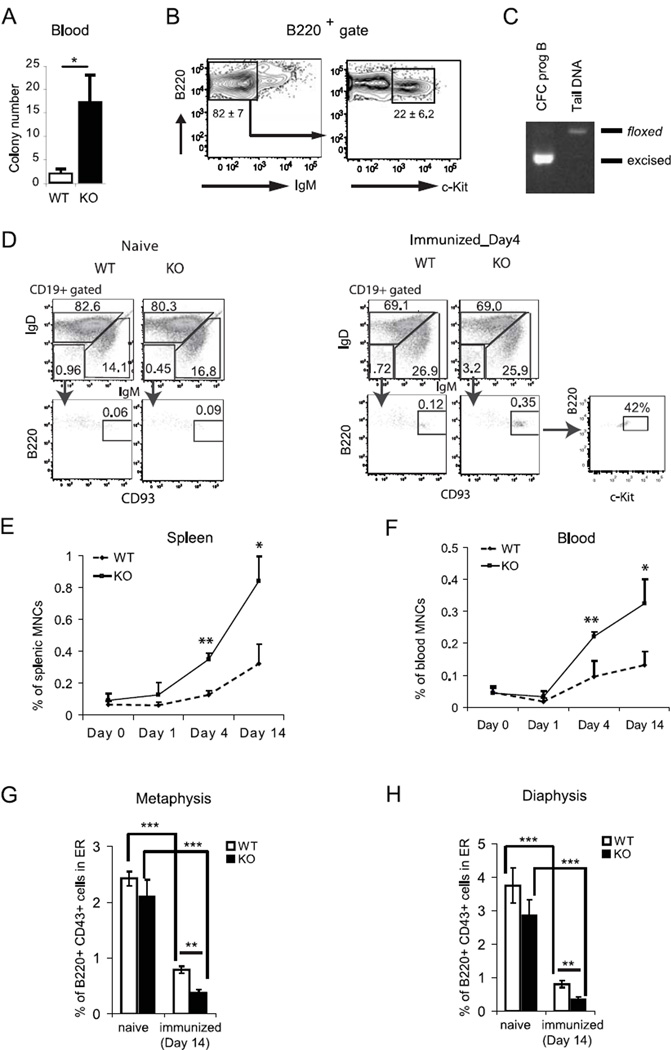
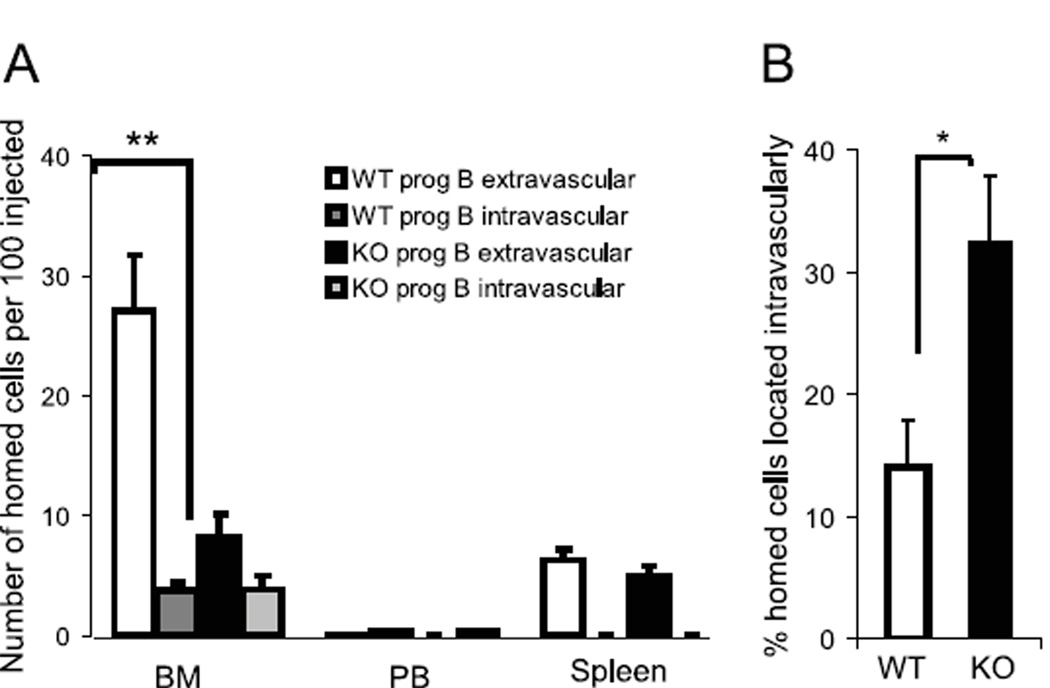
Progenitor B cells reside in complex bone marrow (BM) microenvironments where they receive signals for growth and maturation. We reported previously that the CXCL12-focal adhesion kinase (FAK)-VLA4 pathway plays an important role in progenitor B cell adhesion and migration. In this study, we have conditionally targeted in B cells FAK, and found that the numbers of progenitor pro-B, pre-B, and immature B cells are reduced by 30-40% in B cell-specific FAK knockout mice. When cultured in methylcellulose with IL-7 ± CXCL12, Fak-deleted pro-B cells yield significantly fewer cells and colonies. Using in situ quantitative imaging cytometry, we establish that in longitudinal femoral BM sections, pro-B cells are preferentially localized in close proximity to the endosteum of the metaphyses and the diaphysis. Fak deletion disrupts the nonrandom distribution of pro-B cells and induces the mobilization of pro-B cells to the periphery in vivo. These effects of Fak deletion on pro-B cell mobilization and localization in BM are amplified under inflammatory stress, that is, after immunization with nitrophenol-conjugated chicken γ-globulin in alum. Collectively, these studies suggest the importance of FAK in regulating pro-B cell homeostasis and maintenance of their spatial distribution in BM niches.
Figures
References
-
- Hardy RR, Hayakawa K. B cell development pathways. Annu Rev Immunol. 2001;19:595–621. - PubMed
-
- Nagasawa T. Microenvironmental niches in the bone marrow required for B-cell development. Nat Rev Immunol. 2006;6:107–116. - PubMed
-
- Visnjic D, Kalajzic Z, Rowe DW, Katavic V, LorenzoH J, Aguila HL. Hematopoiesis is severely altered in mice with an induced osteoblast deficiency. Blood. 2004;103:3258–3264. - PubMed
-
- Osmond DG, Kim N, Manoukian R, Phillips RA, Rico-Vargas SA, Jacobsen K. Dynamics and localization of early B-lymphocyte precursor cells (pro-B cells) in the bone marrow of scid mice. Blood. 1992;79:1695–1703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
