

The intramembrane protease SPPL2a promotes B cell development and controls endosomal traffic by cleavage of the invariant chain
- PMID: 23267015
- PMCID: PMC3549707
- DOI: 10.1084/jem.20121069
The intramembrane protease SPPL2a promotes B cell development and controls endosomal traffic by cleavage of the invariant chain
Abstract
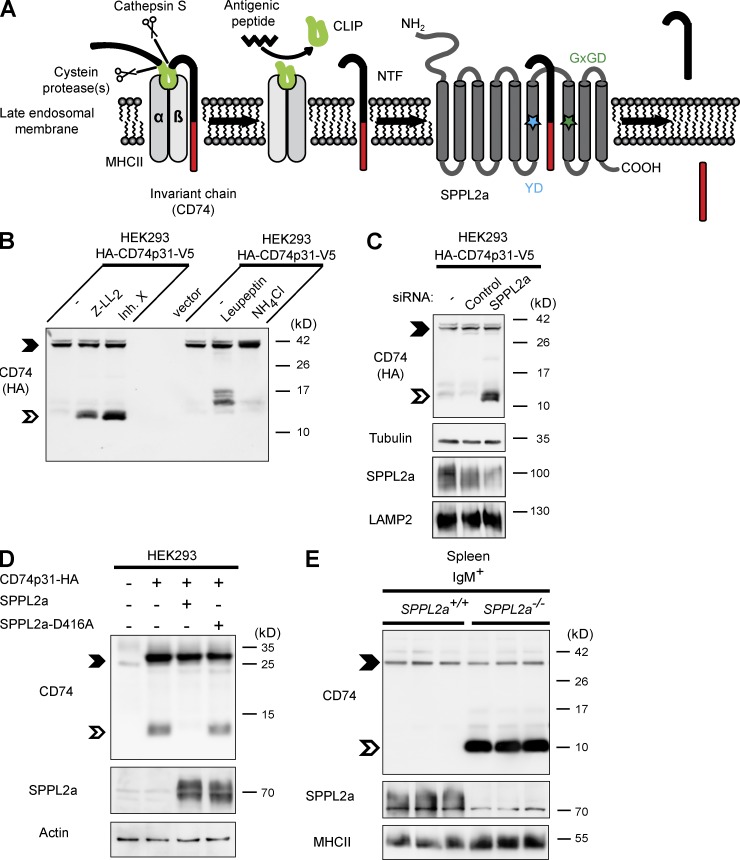
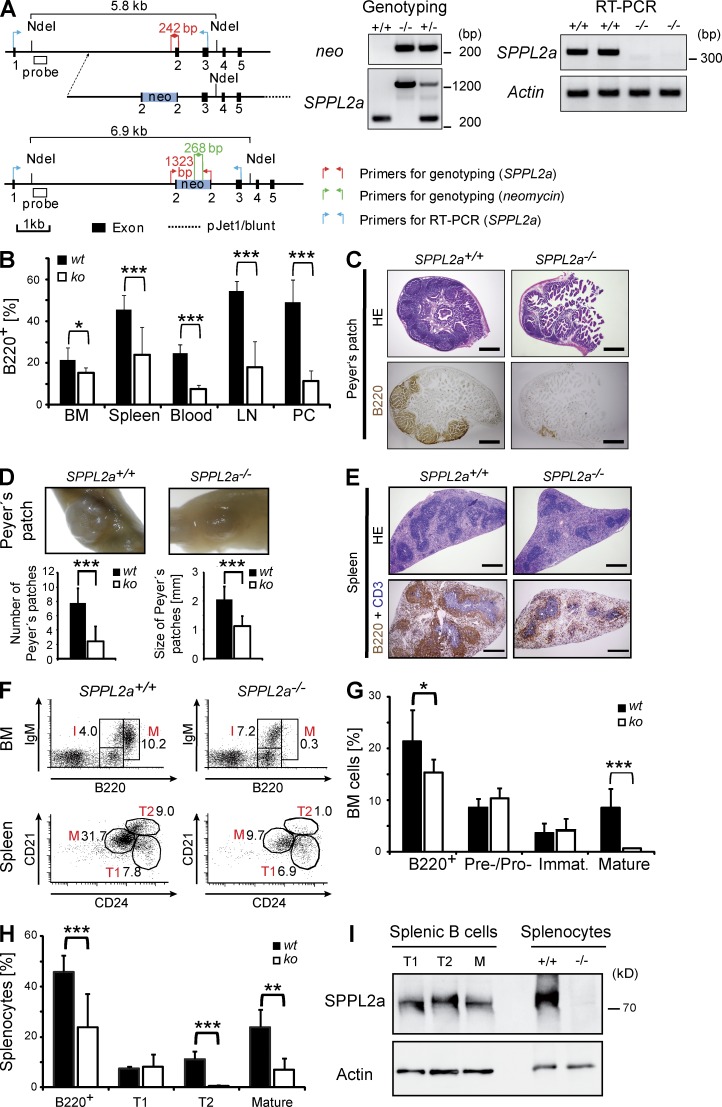
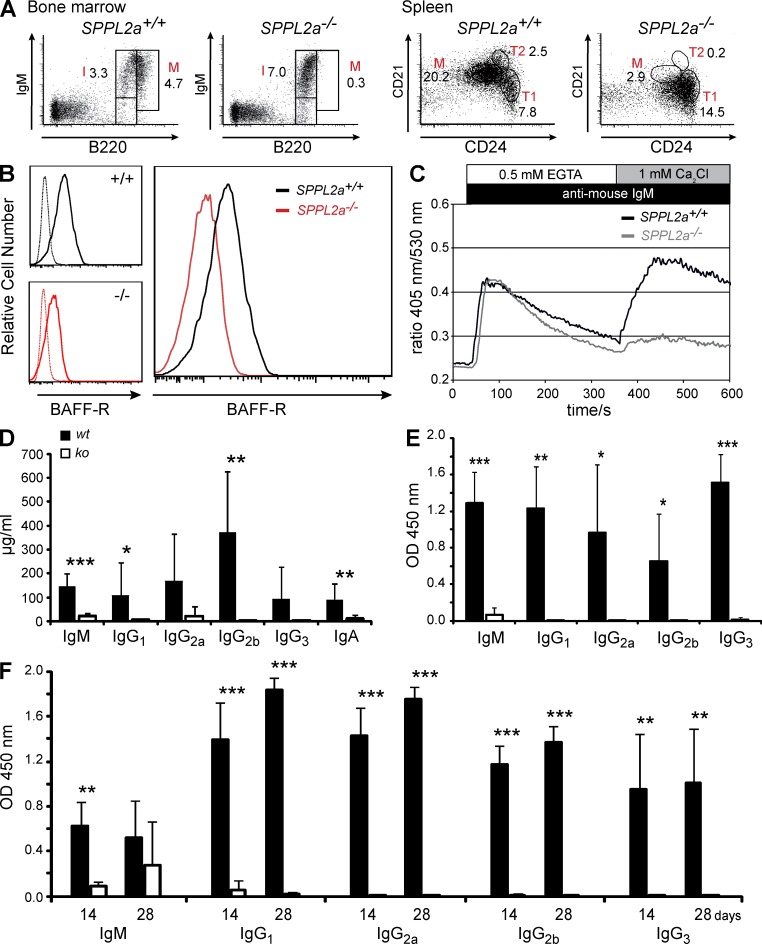
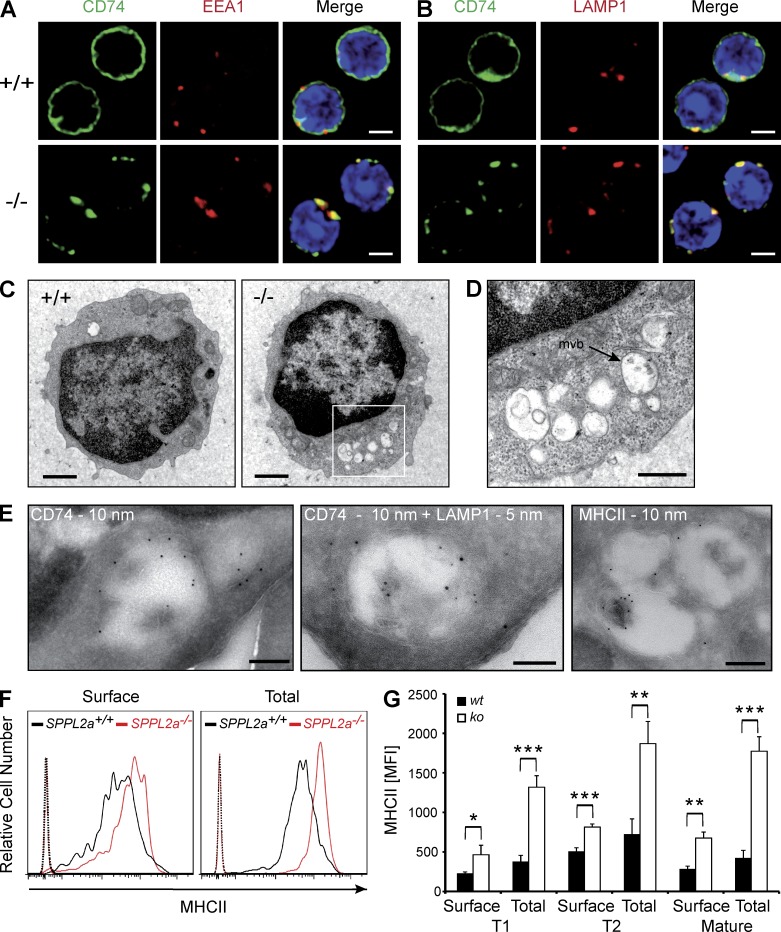
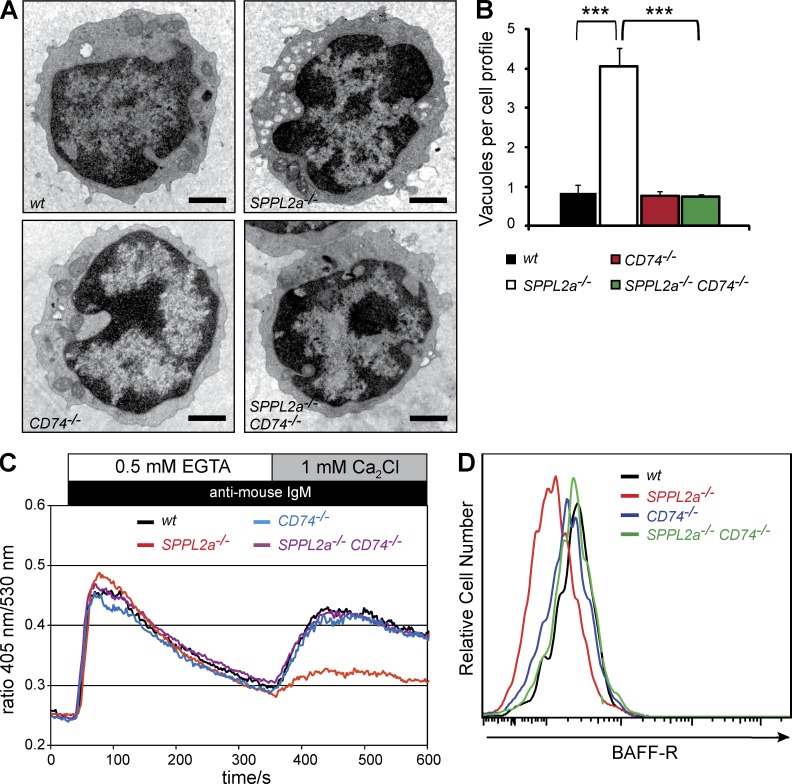
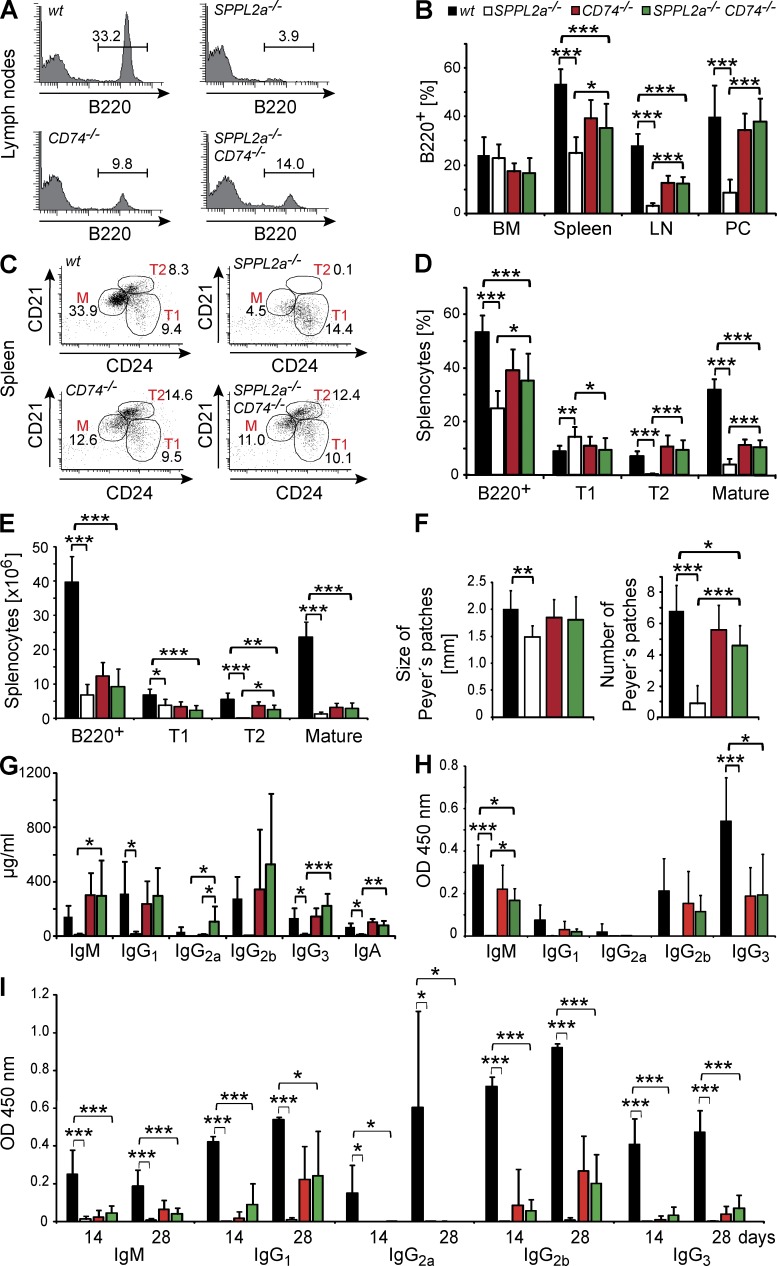
Regulated intramembrane proteolysis is a central cellular process involved in signal transduction and membrane protein turnover. The presenilin homologue signal-peptide-peptidase-like 2a (SPPL2a) has been implicated in the cleavage of type 2 transmembrane proteins. We show that the invariant chain (li, CD74) of the major histocompatability class II complex (MHCII) undergoes intramembrane proteolysis mediated by SPPL2a. B lymphocytes of SPPL2a(-/-) mice accumulate an N-terminal fragment (NTF) of CD74, which severely impairs membrane traffic within the endocytic system and leads to an altered response to B cell receptor stimulation, reduced BAFF-R surface expression, and accumulation of MHCII in transitional developmental stage T1 B cells. This results in significant loss of B cell subsets beyond the T1 stage and disrupted humoral immune responses, which can be recovered by additional ablation of CD74. Hence, we provide evidence that regulation of CD74-NTF levels by SPPL2a is indispensable for B cell development and function by maintaining trafficking and integrity of MHCII-containing endosomes, highlighting SPPL2a as a promising pharmacological target for depleting and/or modulating B cells.
Figures
References
-
- Beisner D.R., Langerak P., Parker A.E., Dahlberg C., Otero F.J., Sutton S.E., Poirot L., Barnes W., Young M.A., Niessen S., et al. 2013. The intramembrane protease Sppl2a is required for B cell and DC development and survival via cleavage of the invariant chain. J. Exp. Med. 210:23–30 - PMC - PubMed
-
- Bergmann H., Yabas M., Short A., Miosge L., Barthel N., Teh C.E., Roots C.M., Bull K., Jeelall Y., Horikawa K., et al. 2012. B cell survival, surface BCR and BAFFR expression, CD74 metabolism, and CD82 dendritic cells require the intramembrane endopeptidase SPPL2A. J. Exp. Med. 210:31–40 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
