

Laccase detoxification mediates the nutritional alliance between leaf-cutting ants and fungus-garden symbionts
- PMID: 23267060
- PMCID: PMC3545816
- DOI: 10.1073/pnas.1212709110
Laccase detoxification mediates the nutritional alliance between leaf-cutting ants and fungus-garden symbionts
Abstract
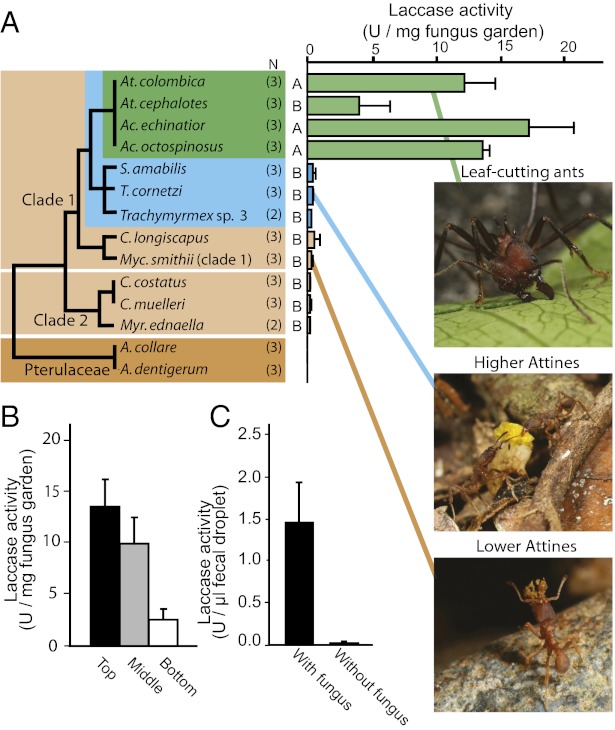
Leaf-cutting ants combine large-scale herbivory with fungus farming to sustain advanced societies. Their stratified colonies are major evolutionary achievements and serious agricultural pests, but the crucial adaptations that allowed this mutualism to become the prime herbivorous component of neotropical ecosystems has remained elusive. Here we show how coevolutionary adaptation of a specific enzyme in the fungal symbiont has helped leaf-cutting ants overcome plant defensive phenolic compounds. We identify nine putative laccase-coding genes in the fungal genome of Leucocoprinus gongylophorus cultivated by the leaf-cutting ant Acromyrmex echinatior. One of these laccases (LgLcc1) is highly expressed in the specialized hyphal tips (gongylidia) that the ants preferentially eat, and we confirm that these ingested laccase molecules pass through the ant guts and remain active when defecated on the leaf pulp that the ants add to their gardens. This accurate deposition ensures that laccase activity is highest where new leaf material enters the fungus garden, but where fungal mycelium is too sparse to produce extracellular enzymes in sufficient quantities to detoxify phenolic compounds. Phylogenetic analysis of LgLcc1 ortholog sequences from symbiotic and free-living fungi revealed significant positive selection in the ancestral lineage that gave rise to the gongylidia-producing symbionts of leaf-cutting ants and their non-leaf-cutting ant sister group. Our results are consistent with fungal preadaptation and subsequent modification of a particular laccase enzyme for the detoxification of secondary plant compounds during the transition to active herbivory in the ancestor of leaf-cutting ants between 8 and 12 Mya.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
The fungal symbiont of Acromyrmex leaf-cutting ants expresses the full spectrum of genes to degrade cellulose and other plant cell wall polysaccharides.BMC Genomics. 2013 Dec 28;14:928. doi: 10.1186/1471-2164-14-928. BMC Genomics. 2013. PMID: 24373541 Free PMC article.
-
Leaf-cutting ant fungi produce cell wall degrading pectinase complexes reminiscent of phytopathogenic fungi.BMC Biol. 2010 Dec 31;8:156. doi: 10.1186/1741-7007-8-156. BMC Biol. 2010. PMID: 21194476 Free PMC article.
-
Ant mediated redistribution of a xyloglucanase enzyme in fungus gardens of Acromyrmex echinatior.BMC Microbiol. 2016 May 6;16:81. doi: 10.1186/s12866-016-0697-4. BMC Microbiol. 2016. PMID: 27154066 Free PMC article.
-
The prominent role of fungi and fungal enzymes in the ant-fungus biomass conversion symbiosis.Appl Microbiol Biotechnol. 2014 Jun;98(11):4839-51. doi: 10.1007/s00253-014-5708-5. Epub 2014 Apr 15. Appl Microbiol Biotechnol. 2014. PMID: 24728757 Review.
-
The origin of the attine ant-fungus mutualism.Q Rev Biol. 2001 Jun;76(2):169-97. doi: 10.1086/393867. Q Rev Biol. 2001. PMID: 11409051 Review.
Cited by
-
Functional characterization of a yellow laccase from Leucoagaricus gongylophorus.Springerplus. 2015 Oct 30;4:654. doi: 10.1186/s40064-015-1464-y. eCollection 2015. Springerplus. 2015. PMID: 26543788 Free PMC article.
-
Examining the Assembly of the Bacterial Community Associated with the Fungus Garden of Atta mexicana (Hymenoptera: Formicidae): Does Queen Origin Matter?Curr Microbiol. 2025 Jul 28;82(9):417. doi: 10.1007/s00284-025-04405-9. Curr Microbiol. 2025. PMID: 40719782
-
The bark beetle holobiont: why microbes matter.J Chem Ecol. 2013 Jul;39(7):989-1002. doi: 10.1007/s10886-013-0318-8. Epub 2013 Jul 12. J Chem Ecol. 2013. PMID: 23846183 Review.
-
Comparative transcriptomic analysis of the mechanisms underpinning ageing and fecundity in social insects.Philos Trans R Soc Lond B Biol Sci. 2021 Apr 26;376(1823):20190728. doi: 10.1098/rstb.2019.0728. Epub 2021 Mar 8. Philos Trans R Soc Lond B Biol Sci. 2021. PMID: 33678016 Free PMC article.
-
The fungal symbiont of Acromyrmex leaf-cutting ants expresses the full spectrum of genes to degrade cellulose and other plant cell wall polysaccharides.BMC Genomics. 2013 Dec 28;14:928. doi: 10.1186/1471-2164-14-928. BMC Genomics. 2013. PMID: 24373541 Free PMC article.
References
-
- Coley PD, Bryant JP, Chapin FS., 3rd Resource availability and plant antiherbivore defense. Science. 1985;230(4728):895–899. - PubMed
-
- Shipley LA, Forbey JS, Moore BD. Revisiting the dietary niche: When is a mammalian herbivore a specialist? Integr Comp Biol. 2009;49(3):274–290. - PubMed
-
- Berryman AA. What causes population cycles of forest Lepidoptera? Trends Ecol Evol. 1996;11(1):28–32. - PubMed
-
- Wirth R, Herz H, Ryel RJ, Beyschlag W, Hölldobler B. 2003. Herbivory of Leaf-Cutting Ants: A Case Study on Atta colombica in the Tropical Rainforest of Panama, eds Baldwin IT, et al., Ecological studies (Springer, Berlin), Vol. 164.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
